
Классификация ВПР
Согласно международной классификации (ВОЗ) врожденные пороки развития подразделяют:
1) по частоте: на распространенные – 1: 1000 и более, умеренно частые –
0,1-0,99 на -1000 и очень редкие – меньше 0,01 на 1000;
2) по степени тяжести и прогнозу для жизнеспособности: на летальные – смерть до репродуктивного периода (0,6%), средней степени тяжести – не угрожают жизни, но требуют оперативного вмешательства (1,9-2,5%) и
малые аномалии развития или информативные морфологические варианты
(3,5%); 3) по проявлению: на изолированные (монотопический дефект поля,
политопический дефект поля и sequence), множественные (синдром,
ассоциация, случайныекомбинации);
4) по этиологии: на моногенные – 6%, хромосомные –5%, внешнесредовые
(тератогены, материнские факторы) – 6%, мультифакториальные – 63%,
неустановленные причины –20%.
В зависимости от последовательности возникновения (типа «ошибок») в
процессе морфогенеза пороки развития делят на 4 группы (Опитц Дж., 1977,
Лазюк Г.И., 1983, Козлова С.И., 1986):
—мальформация (malformation) – морфологический дефект органа, части органа или большого участка тела в результате внутреннего нарушения процесса развития (генетически детерминированный процесс);
—дизрупция (disruption)- морфологический дефект органа, части органа или большого участка тела в результате внешнего препятствия или какого-либо воздействия на изначально нормальный процесс развития (тератогенные факторы и нарушение имплантации);
—деформация (deformation) –нарушение формы, вида или положения части тела, обусловленное механическими воздействиями;
—дисплазия (dysplasia)- нарушенная организация клеток в ткани (тканях) и ее морфологический результат (процесс и следствие дисгистогенеза).
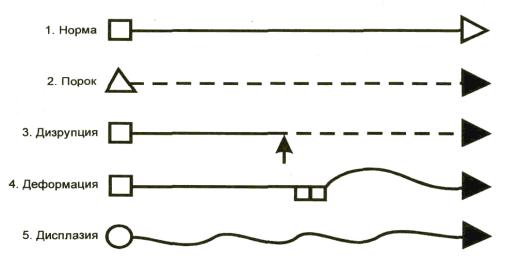
Рис. 9. Схема разных типов ошибок морфогенеза (J. Opitz, 1977).
В зависимости от времени и объекта воздействия вредных факторов ВПР делятся на четыре типа: гаметопатии – связаны с возникновением мутаций в половых клетках родителей (генные, хромосомные синдромы);
бластопатии – поражение бластоцисты (зародыш первых 15 дней после оплодотворения, следствие бластопатий – двойниковые пороки, циклопия,
сиреномелия, пустые зародышевые мешки); эмбриопатии – возникают в период от 16 дня до конца 8 недели беременности (в этот период происходит активный органогенез и гистогенез, большинство пороков образуется в этот период); фетопатии – повреждение плода в период от 9 недели до окончания беременности (крипторхизм, тазовая почка, метаболические фетопатии). Ниже мы приводим описание наиболее известных эмбриофетопатий.
Фетальный вальпроевый синдром
Данный синдром обусловлен приемом во время беременности противосудорожных препаратов вальпроевой кислоты. Дети рождаются с дефицитом массы и длины тела. Типичны аномалии лица: небольшие орбиты, гипоплазия средней части лица, короткий вздернутый нос, длинный уплощенный фильтр, широкая переносица, эпикант, микрогнатия, маленький рот, расщелина губы, расщелина неба, микротия, анотия. Также для
синдрома характерны аномалии черепа: тригоноцефалия, узкий бифронтальный диаметр, синостоз швов. К другим проявлениям синдрома относятся полидактилия, дефекты нервной трубки (спинномозговая грыжа),
пороки сердца (ДМЖП, ОАП, коарктация аорты), гипоспадия, ВПР ЖКТ. У
детей наблюдается отставание в статико-моторном и умственном развитии.
Фетальный синдром краснухи (синдром Грега)
Впервые синдром описан в 1941 году N. Gregg. Риск для плода у серопозитивных женщин составляет 40-90%. Инфицирование плода вирусом краснухи может вызвать спонтанный аборт, преждевременные роды.
Типичны поражения сердца, органов зрения и слуха. Среди ВПР глаз отмечаются катаракта, микрофтальмия, пигментная ретинопатия, глаукома,
миопия, страбизм, колобома. Постоянный признак – нейросенсорная глухота
(одноили двусторонняя). Из ВПР сердца встречаются открытый аортальный проток, стеноз легочной артерии, дефект межжелудочковой перегородки. Характерно выделение вируса из различных тканей, выявление специфических антител в сыворотке плода. У больных детей наблюдается умственная отсталость, дефицит роста, микроцефалия, гидроцефалия,
пренатальная гипоплазия, иммунологический дефицит, персистенция вируса во многих органах, аномалии зубов, гемолитическая анемия. Основным видом профилактики является иммунизация девочек по общепринятой схеме.
Фетальный цитомегаловирусный синдром
Высокий риск развития данного синдрома связан с первичной цитомегаловирусной инфекцией в первые 6 месяцев гестации. У 10% плодов имеются тяжелые ВПР. Характерны внутриутробная задержка развития,
гепатоспленомегалия, желтуха, инфицирование плода цитомегаловирусом.
Дети рождаются недоношенными, с низкой массой тела. Нередко отмечаются микроцефалия, перивентрикулярные кальцификаты (энцефалит),
гидроцефалия, умственная отсталость, гипотония, судороги. Возможно поражение глаз: страбизм, хореоретинит, микрофтальмия, катаракта,
кальцификаты в сетчатке. Часто у детей выявляется тромбоцитопения,
гемолитическая анемия, петехии. Профилактика: избегать контактов,
пренатальная диагностика (УЗИ) – вентрикуломегалия, кальцификаты в головном мозге.
Фетальный алкогольный синдром
(FAS, алкогольная эмбриофетопатия)
Частота ФАС в США — от 0,2 до 2 на 1000. При хроническом алкоголизме матери риск развития ФАС составляет 25-45%. У детей отмечается дефицит роста и веса. Характерно поражение центральной нервной системы: тремор,
судороги, опистотонус, гипотония, нарушения поведения, умственная отсталость, гидроцефалия, микроцефалия. Типичны изменения лица:
короткая глазная щель, сглаженный губной желобок, тонкая верхняя губа,
блефарофимоз, эпикант, птоз, косоглазие, удлиненное лицо, микрогения. У
детей выявляются аномалии пальцев, дисплазия ТБС, деформации грудной клетки, гипоспадия, крипторхизм, удвоение влагалища, ВПР сердца (чаще ДМЖП), ангиомы, фиброз печени.
В медицинской практике также широко используется классификация аномалий развития, которая основана на органно-системном (анатомо-
физиологическом) принципе.
К распространенным видам пороков в практике врача любой специальности относятся следующие:
—агенезия – полное врожденное отсутствие органа;
—аплазия – врожденное отсутствие органа с сохранением его сосудистой ножки;
—атрезия и стеноз – соответственно полное отсутствие или сужение канала
(отверстия);
— врожденная гипоплазия – недоразвитие органа, проявляющееся дефицитом относительной массы (отношение абсолютной массы органа к абсолютной массе тела плода или ребенка, выраженное в процентах) или размеров органа, превышающее отклонение от средних показателей в две сигмы
(средняя ошибка) для данного возраста: простая – без нарушения структуры и диспластическая – с нарушением структуры;
— врожденная гипотрофия – уменьшение массы тела плода или новорож-
денного;
— врожденная гипертрофии (гиперплазия) – увеличение относительной мас-
сы (размеров) органа за счет увеличения объема (гипертрофия) или количества (гиперплазия) клеток;
— гетероплазия – нарушение дифференцировки отдельных типов тканей,
например, клетки плоского эпителия пищевода в дивертикуле Меккеля;
—гетеротопия – наличие клеток, тканей или участка органа в другом органе или в тех зонах органа, где в норме их не должно быть;
—эктопия – смещение органа в нетипичное для него место;
—дизрафия или арафия – незаращение эмбриональной щели (расщелина губы или неба, позвоночника);
—дисхрония – нарушение темпов развития (ускоренное или замедленное);
—макросомия (гигантизм) – увеличение длины тела;
—неразделение (слияние) – может быть органа или монозиготных близнецов
(краниопаги, торокопаги, сросшиеся черепами или грудными клетками близнецы);
— персистирование – сохранение эмбриональных структур, в норме исчеза-
ющих к определенному периоду онтогенеза, например, незаращение баталлова протока или овального окна у ребенка в возрасте старше трех месяце;
— нарушение лобуляции – дополнительная доля легкого, печени;
—инверсия-зеркальноерасположениеорганов:
—удвоение органов;
—образование водянок – гидроцефалия, гидронефроз.
Для объяснения этиологических и патогенетических связей между разными типами аномалий развития предложены следующие понятия:
следствие, синдром, ассоциация.
Синдром – комплекс множественных пороков и аномалий развития,
патогенетически и этиологически связанных между собой; это устойчивое сочетание двух и более не индуцированных друг другом пороков развития в разных системах организма.
Следствие (аномалад, секвенция, вторичный порок) – комплекс пороков развития, возникающий вследствие действия одного ВПР или физического фактора, который вызывает каскад вторичных нарушений (связь между пороками патогенетическая). Например, спорадический аномалад Пьера-
Робена, при котором первичным пороком является микрогения, а его следствием будут уменьшение ротовой полости, глоссоптоз, расщелина неба.
Ассоциация – неслучайное сочетание нескольких пороков и аномалий развития, наблюдающееся у 2-х и более пациентов, не известных как синдром или следствие. Например, ассоциация OEIS – омфалоцеле,
экстрофия мочевого пузыря, атрезия заднего прохода, дефекты позвоночника; ассоциация VAСTERL (MIM:192350) – атрезия пищевода и заднего прохода, трахео-эзофагальные фистулы, пороки почек, сердца,
дефекты позвоночника, радиальные дисплазии.
В последнее время внимание врачей различных специальностей обращается на диагностическое значение малых аномалий развития. Малая аномалия развития (МАР, дисгенезия, информативный морфологический вариант, дизэмбриологический признак, дисморфологическая черта,
микропризнак, микродегенеративный признак и др.) – это стойкое морфологическое нарушение органа, участка тела, не выходящее за пределы вариаций или находящееся у крайних границ вариаций строения и не сопровождающееся нарушением его функции. Известно около 200 МАР.
Малые аномалии развития с разной частотой распространены в популяции в рамках нормальной вариабельности. Например, сросшиеся брови,
поперечная складка ладони, гипертелоризм, деформация ушных раковин,
клинодактилия V пальца, гипертрихоз и др. Микропризнаки – это морфологические нарушения на конечной тонко настроенной стадии
развития, т.е. нарушения гистогенеза. Для них характерна стабильность – имеются при рождении, не исчезают с возрастом (исключение капиллярная гемангиома, эпикант, пигментные пятна и др.). Малые аномалии не имеют серьезного медицинского или косметического значения, но нередко выступают как значащие симптомы наследственной патологии. Выявление данных изменений фенотипа ребенка требует динамического наблюдения за ним и проведения цитогенетического и ультразвукового исследований.
В диагностических целях следует учитывать 3 важных параметра МАР: их количество, сочетание, «качество». При наличии 3-5 и более микропризнаков
часто выявляются большие аномалии развития или нарушения интеллекта. В
качестве маркеров некоторых наследственных заболеваний и синдромов МВПР могут рассматриваться комплексы из 4, 5-и и более МАР. Например,
полидактилия, гипертелоризм глаз и сосков, дополнительные соски – эти
«малые» аномалии предполагают наличие у пациента пороков мочевыводящей системы. Большое значение имеет сочетание МАР.
Например, для синдрома Дауна характерно наличие монголоидного разреза глаз, эпиканта, брахидактилии, клинодактилии, сандалевидной щели,
поперечной ладонной складки. Наибольшее диагностическое значение имеет
«качество» МАР, т.е. выделение таких признаков, которые среди здоровых пациентов почти не встречаются: «крыловидные» складки шеи
(с.Шерешевского-Тернера, с.Нунан), постаксиальная полидактилия (с.Барде-
Бидля), вертикальные насечки на мочке уха (с.Беквита-Видемана).
Наибольшей значимостью обладают такие микропризнаки, которые встречаются редко и среди больных с наследственной и врожденной патологией, и в популяции здоровых людей. Например, диастема (частота в популяции 15%) и гиподонтия (очень редко встречающийся признак) несут разную диагностическую нагрузку. Отдельные микропризнаки встречаются чаще у недоношенных и маловесных детей. Они могут быть этническими особенностями, например, эпикант является нормой у монголоидной расы –
60-65%. МАР можно разделить на 3 группы: альтернативные, измерительные
1.2. Проблемы морфогенеза
Биологический морфогенез можно определить как «появление характерной и специфической формы в живых организмах».[18] Первая проблема есть именно та, что форма обретает существование. Биологическое развитие эпигенетическое: появляются новые структуры, которые не могут быть объяснены как результат развертывания или роста структур, которые уже присутствуют в яйце в начале развития.
Вторая проблема состоит в том, что многие развивающиеся системы способны регулировать, другими словами, если часть развивающейся системы удаляется (или если добавляется дополнительная часть), система продолжает развиваться таким образом, что образуется более или менее нормальная структура. Классическая демонстрация этого явления была проведена в 1890-е годы Г. Дришем в его экспериментах на эмбрионах морского ежа. Когда одну из клеток очень молодого эмбриона на двухклеточной стадии развития убивали, из оставшейся клетки развивалась не половина морского ежа, но совершенно целый морской еж, хотя и меньшего размера. Точно так же маленькие, но целые организмы развивались после разрушения любых одной, двух или трех клеток эмбриона на четырех-клеточной стадии. Напротив, после слияния двух молодых эмбрионов морского ежа развивался один гигантский морской еж.[19]
Регуляция была продемонстрирована на многих развивающихся системах. Однако в процессе развития организма эта способность утрачивается, когда определяется судьба его отдельных частей. Но даже в системах, где эта детерминированность имеет место на ранней стадии, например в эмбрионах насекомых, регуляция может осуществляться после повреждения яйца (рис. 1).
Результаты такого типа показывают, что развивающиеся системы движутся к морфологической цели и что они обладают некоторым свойством, которое определяет эту цель и позволяет им достичь ее, даже если части системы удалены и созданы препятствия для нормального хода развития.
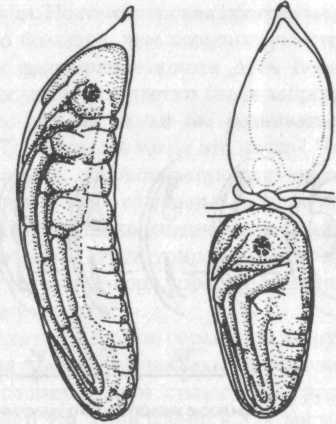
Рис. 1. Пример регуляции. Слева — нормальный эмбрион стрекозы Platycnemis pennipes. Справа — небольшой, но полноценный эмбрион, полученный из задней части яйца, перетянутого в середине вскоре после его откладывания (из Weiss, 1942)
Третья проблема — это регенерация, то есть способность организмов заменять или восстанавливать поврежденные структуры. Растения обнаруживают удивительно широкий диапазон возможностей регенерации, и то же можно сказать о низших животных:
например, если плоского червя разрезать на несколько кусочков, из каждого может вырасти целый червь. Даже многие позвоночные обладают поразительными способностями к регенерации: например, если хирургическим путем удалить линзу из глаза тритона, из края радужной оболочки вырастает новая линза (рис. 2); при нормальном эмбриональном развитии линза образуется совершенно другим путем, из клеток кожи. Этот тип регенерации был впервые обнаружен Г. Вольфом. Он умышленно выбрал такой вид повреждения организма, который не мог случайно произойти в природе; поэтому такой регенеративный процесс не мог быть предметом естественного отбора.[20]
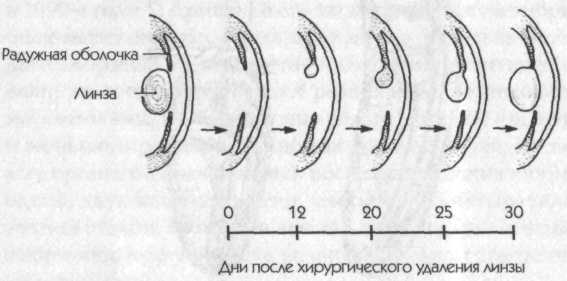
Рис. 2. Регенерация линз из границ радужной оболочки глаза тритона после хирургического удаления природной линзы (Needham, 1942)
Четвертая проблема — это просто сам факт воспроизведения: отдельная часть родительского организма становится новым организмом, часть становится целым.
Эти явления могут быть понятны, только если допустить наличие причинных сущностей, которые составляют нечто большее, нежели сумма частей развивающихся систем, и определяют цели процессов развития.
Виталисты приписывают эти свойства витальным факторам, органицисты — морфогенетическим полям, механицисты — генетическим программам.
Концепция генетических программ основана на аналогии с программами, определяющими работу компьютеров. Она предполагает, что оплодотворенное яйцо содержит предварительно сформированную программу, которая каким-то образом определяет морфогенетические задачи организма, а также координирует и контролирует его развитие в направлении, соответствующем этим задачам. Но генетическая программа должна включать нечто большее, чем химическую структуру ДНК, поскольку идентичные копии ДНК передаются всем клеткам; если бы все клетки были запрограммированы одинаково, они не могли бы развиваться различным образом. Тогда что же такое это нечто? Чтобы ответить на этот вопрос, основополагающую идею приходится представлять в виде отдельных неопределенных предположений о физико-химических взаимодействиях, каким-то образом структурированных во времени и пространстве; иначе говоря, проблема лишь предстает в иной формулировке.[21]
Существует и другое серьезное затруднение. Компьютерная программа закладывается в компьютер разумным сознательным существом, программистом. Она создается и записывается для достижения определенной цели. Если генетическая программа рассматривается как аналог компьютерной программы, то тогда должна существовать некая целенаправленно действующая сущность, исполняющая роль программиста. Но если предположить, что генетические программы аналогичны не обычным компьютерным программам, а программам самовоспроизводящихся, самоорганизующихся компьютеров, тогда проблема состоит в том, что таких компьютеров просто не существует. А если бы даже они существовали, то должны были бы быть невероятно сложным образом запрограммированы теми, кто их изобрел. Единственный выход из этой дилеммы — утверждать, что генетическая программа построена в ходе эволюции путем сочетания случайных мутаций и естественного отбора. Но тогда подобие с компьютерной программой исчезает и эта аналогия становится бессмысленной.
Ортодоксальные механицисты отвергают идею, что кажущееся целенаправленным поведение развивающихся, регулирующих и регенерирующих организмов указывает на присутствие витального фактора, который ведет их к достижению морфологических целей. Но до тех пор, пока механистические объяснения зависят от телеологических концепций, таких как генетические программы или генетические инструкции, целенаправленность может быть объяснена только тем, что она уже была заложена в них ранее. Действительно, свойства, приписываемые генетическим программам, удивительно похожи на те, которыми виталисты наделяют свои гипотетические витальные факторы; ирония в том, что генетические программы оказываются весьма похожими на витальный фактор в механистическом облике.[22]
Конечно, тот факт, что биологический морфогенез в настоящее время не может быть объяснен в строго механистическом духе, не доказывает, что он не может быть так объяснен никогда. Перспективы получения такого объяснения рассматриваются в следующей главе. Но на сегодняшний день убедительный ответ с механистических позиций дать невозможно.
Читайте также
Лесная нива и ее проблемы
Лесная нива и ее проблемы
Постановка проблемы
Постановка проблемы
Четыре ноги хорошо, две ноги — плохо.
Дж. Оруэлл «Скотный двор»
Способность к счету является, несомненно, одной из высших психических функций живых организмов. Вопрос о том, в чем заключается понятие «истинного счета» и действительно ли некоторые
§ 18. Биологические проблемы мышления
§ 18. Биологические проблемы мышления
Обладатели крупного мозга становятся своеобразными заложниками его размеров. Нравится или не нравится животному, но в большом количестве нейрональных связей постоянно фиксируется повседневная информация о внешнем мире и
Проблемы общения
Проблемы общения
В концепции Л.В. Крушинского о рассудочной деятельности интересно представление о двух его механизмах, особенно о вербальном. Автор допускает существование вербального мышления у животных, в частности у человекообразных обезьян, и возможность
Проблемы макроэволюции
Проблемы макроэволюции
В рамках макроэволюции рассматривается широкий круг вопросов, касающихся закономерностей эволюции таксонов. Это анализ филогенетических изменений органов в связи с их функцией, анализ основных типов филогенеза и факторов, их определяющих,
БИОЛОГИЧЕСКИЕ АСПЕКТЫ ПРОБЛЕМЫ
БИОЛОГИЧЕСКИЕ АСПЕКТЫ ПРОБЛЕМЫ
Термитов часто называют белыми муравьями, но этот термин может ввести в заблуждение. В действительности термиты ведут свою родословную от тараканов, которые, в свою очередь, появились на Земле свыше 200 миллионов лет назад, задолго до других
Пространственная организация и начало морфогенеза
Пространственная организация и начало морфогенеза
Сложные изменения размеров и формы, которые составляют морфологическую эволюцию, могут происходить довольно быстро и достигаются главным образом в результате изменений не структурных генов, а регуляторных элементов,
Глава 2. Три теории морфогенеза
Глава 2. Три теории морфогенеза
2.1. Описательное и экспериментальное исследование
Описание развития можно проводить различными путями: внешняя форма развивающегося животного или растения может быть зарисована, сфотографирована или снята на кинопленку, давая картину
6.4. Возрастающая специфичность морфического резонанса в процессе морфогенеза
6.4. Возрастающая специфичность морфического резонанса в процессе морфогенеза
Энергетический резонанс не является процессом типа «все или ничего»: система резонирует в ответ на диапазон частот, более или менее близких к ее естественной частоте, хотя максимальный отклик
7.3. Измененные пути морфогенеза
7.3. Измененные пути морфогенеза
В то время как факторы, влияющие на морфогенетические зародыши, производят в морфогенезе качественные эффекты, такие как отсутствие какой-либо структуры или замена одной структуры на другую, многие генетические факторы или воздействия
Проблемы
Проблемы
Современные
проблемы — экологические, политические, социальные, бытовые — в конечном счете
представляют собой различные аспекты конфликтности человеческого существования,
в основе которой лежит конкуренция между отдельными людьми, экономическими
7.3. Другие проблемы медицины
7.3. Другие проблемы медицины
Концепция универсальных функциональных блоков существенно влияет на понимание патологии, геронтологии, фармакологической регуляции различных видов стресса и т.д. Достаточно вспомнить, что С-концевые фрагменты гастрина и холецистокинина
Проблемы стока
Проблемы стока
Большое значение для охраны вод суши имеют Основы водного законодательства Союза ССР и союзных республик, которые были приняты Верховным Советом СССР в декабре 1970 г. В законе подчеркивается, что в результате Великой Октябрьской социалистической
Схема разных типов ошибок морфогенеза — Справочник по медицине PRO7
Review
. 1998 Apr 1;76(4):291-6.
Affiliations
-
PMID:
9545092
Review
Errors of morphogenesis and developmental field theory
M L Martínez-Frías et al.
Am J Med Genet.
1998.
Abstract
Field theory provides a rational basis for birth defects terminology. During blastogenesis in higher metazoa, pattern formation in the primary field leads to the establishment of upstream expression domains of growth and transcription factors, which, in various permutations and at specific sites and times, lay down the pattern of progenitor fields. Further spatially coordinated, temporally synchronized, and epimorphically hierarchical morphogenetic events, mostly during organogenesis, lead to the attainment of final form in the secondary, epimorphic fields. Because of shared molecular determinants, spatial contiguity, and close timing of morphogenetic events during blastogenesis, most malformations arising during blastogenesis are polytopic, i.e., involving two or more progenitor fields, e.g., acrorenal, cardiomelic, gastromelic, or splenomelic anomalies. Defects of organogenesis tend to be monotopic malformations, e.g., cleft palate or postaxial polydactyly. We suggest that what were called «associations» (e.g., VATER, schisis) be designated primary polytopic developmental field defects, or simply polytopic field defects, and that the term «association» be reserved for the original definition of a statistical combination of anomalies (mostly of organogenesis) [Spranger et al. (1982): J Pediatr 100:160-165]. If genetically caused or predisposed, all structures involved in a polytopic or monotopic malformation are genetically abnormal, whereas the parts secondarily affected as a consequence of a malformation sequence (e.g., spina bifida) are genetically normal. Polytopic field anomalies, per se, must be distinguished from pleiotropy, although such anomalies may constitute a part of pleiotropy (e.g., in trisomy 18). Because they are downstream from pattern-forming events in the primary field, multiple anomalies of organogenesis more likely represent syndromal pleiotropy.
Similar articles
-
[Developmental abnormalities in humans].
Opitz JM.
Opitz JM.
Monatsschr Kinderheilkd. 1991 May;139(5):259-72.
Monatsschr Kinderheilkd. 1991.PMID: 1870596
Review.
German. -
Limb anomalies from evolutionary, developmental, and genetic perspectives.
Opitz JM.
Opitz JM.
Birth Defects Orig Artic Ser. 1996;30(1):35-77.
Birth Defects Orig Artic Ser. 1996.PMID: 9125339
Review.
-
Pathogenetic classification of a series of 27,145 consecutive infants with congenital defects.
Martínez-Frías ML, Bermejo E, Frías JL.
Martínez-Frías ML, et al.
Am J Med Genet. 2000 Jan 31;90(3):246-9.
Am J Med Genet. 2000.PMID: 10678664
Review.
-
Primary developmental field. III: Clinical and epidemiological study of blastogenetic anomalies and their relationship to different MCA patterns.
Martínez-Frías ML, Frías JL.
Martínez-Frías ML, et al.
Am J Med Genet. 1997 May 2;70(1):11-5. doi: 10.1002/(sici)1096-8628(19970502)70:1<11::aid-ajmg3>3.0.co;2-u.
Am J Med Genet. 1997.PMID: 9129734
-
The developmental field concept.
Opitz JM.
Opitz JM.
Am J Med Genet. 1985 May;21(1):1-11. doi: 10.1002/ajmg.1320210102.
Am J Med Genet. 1985.PMID: 4003434
Cited by
-
Patterns of co-occurring birth defects in children with anotia and microtia.
Schraw JM, Benjamin RH, Shumate CJ, Canfield MA, Scott DA, McLean SD, Northrup H, Scheuerle AE, Schaaf CP, Ray JW, Chen H, Agopian AJ, Lupo PJ.
Schraw JM, et al.
Am J Med Genet A. 2023 Mar;191(3):805-812. doi: 10.1002/ajmg.a.63081. Epub 2022 Dec 21.
Am J Med Genet A. 2023.PMID: 36541232
-
Anorectal Malformations in Male Monozygotic Twins and Review of Literature.
Kundal VK, Kundal R, Meena AK, Debnath PR, Shukla AK.
Kundal VK, et al.
J Indian Assoc Pediatr Surg. 2022 Mar-Apr;27(2):136-139. doi: 10.4103/jiaps.JIAPS_344_20. Epub 2022 Mar 1.
J Indian Assoc Pediatr Surg. 2022.PMID: 35937120
Free PMC article. -
Five-year experience of anorectal malformation with oesophageal atresia in tertiary care hospital.
Chowdhary S, Panigrahi P, Kumar R.
Chowdhary S, et al.
Afr J Paediatr Surg. 2020 Jul-Dec;17(3 & 4):49-53. doi: 10.4103/ajps.AJPS_54_17.
Afr J Paediatr Surg. 2020.PMID: 33342833
Free PMC article. -
Maternal drug use and the risk of anorectal malformations: systematic review and meta-analysis.
Zwink N, Jenetzky E.
Zwink N, et al.
Orphanet J Rare Dis. 2018 May 10;13(1):75. doi: 10.1186/s13023-018-0789-3.
Orphanet J Rare Dis. 2018.PMID: 29747656
Free PMC article. -
Full Spectrum of VACTERL in New Born.
Harjai MM, Holla RG, Kale R.
Harjai MM, et al.
Med J Armed Forces India. 2008 Jan;64(1):84-5. doi: 10.1016/S0377-1237(08)80163-3. Epub 2011 Jul 21.
Med J Armed Forces India. 2008.PMID: 27408093
Free PMC article.No abstract available.
Publication types
MeSH terms
LinkOut — more resources
-
Full Text Sources
- Wiley
-
Medical
- MedlinePlus Health Information
Подборка по базе: Ответы на вопросы.docx, Контрольные вопросы к защите.docx, Экзаменационные вопросы.doc, кр вопросы 3.pdf, Тестовые вопросы к разделу 3 просмотр попытки.pdf, контрольные вопросы по истории.docx, педиатрия лечебный факультет вопросы к экзамену-2.doc, 4 класс. Вопросы.docx, РУБЕЖКА2 ИКТ ВОПРОСЫ.docx, 5. Вопросы для самопроверки.docx
Радиационный и химический мутагенез. Стабильность генетического материала и спонтанный мутационный процесс. Механизм индуцированного мутагенеза и его связь с репарацией.
Для получения индуцированных мутаций у растений используют физические мутагены (гамма-излучение, рентгеновское и ультрафиолетовое излучение) и специально созданные химические супермутагены (например, N-метил-N-нитрозомочевина). Дозу мутагенов подбирают таким образом, чтобы погибало не более 30…50% обработанных объектов. Выделение мутантных организмов начинают со второго поколения. Этим путем получены высокопродуктивные штаммы микроорганизмов (продуцентов антибиотиков), карликовые сорта растений с повышенной скороспелостью и т. д. Однако применение индуцированных мутаций все же ограничено, поскольку у животных мутации практически всегда приводят к снижению жизнеспособности и/или бесплодию. К немногим исключениям относится тутовый шелкопряд, с которым велась интенсивная селекционная работа с использованием авто- и аллополиплоидов
- Мейоз, характеристика его стадий. Кроссинговер и его биологический смысл.
Мейоз – это особый способ деления эукариотических клеток, при котором исходное число хромосом уменьшается в два раза. Мейоз сопровождает образование гамет у животных и формированием спор у растений. В результате образуются гаплоидные половые клетки, при слиянии которых во время оплодотворения восстанавливается диплоидный набор хромосом зиготы. Мейоз включает два мейотических деления. А репликация ДНК происходит только один раз.
Главной особенностью мейоза является конъюгация. В первом делении происходит уменьшение числа хромосом в два раза, поэтому его называют редукционным. Во втором делении число хромосом не изменяется; поэтому его называют эквационным (уравнивающим).
Предмейотическая интерфаза отличается от обычной интерфазы тем, что процесс репликации ДНК не доходит до конца: примерно 0,2…0,4 % ДНК остается неудвоенной. Однако в целом, можно считать, что в диплоидной клетке (2n) содержание ДНК составляет 4с. При наличии центриолей происходит их удвоение. В клетке имеется две диплосомы, каждая из которых содержит пару центриолей.
Первое деление мейоза (редукционное или мейоз I)
Профаза I (профаза первого деления) включает ряд стадий.
- Лептотена (стадия тонких нитей). Хромосомы видны в микроскоп в виде клубка тонких нитей.
- Зиготена (стадия сливающихся нитей). Происходит конъюгация гомологичных хромосом, образуются биваленты. Гомологи удерживаются друг около друга с помощью белковых синаптонемальных комплексов. Количество бивалентов равно n. Иначе биваленты называются тетрады.
- Пахитена (стадия толстых нитей). Хромосомы спирализуются, хорошо видна их продольная неоднородность. Завершается репликация ДНК. Завершается кроссинговер – перекрест хромосом, в результате которого они обмениваются участками хроматид.
- Диплотена (стадия двойных нитей). Гомологичные хромосомы в бивалентах отталкиваются друг от друга. Они соединены в отдельных точках, которые называются хиазмы.
- Диакинез (стадия расхождения бивалентов). Хиазмы перемещаются к теломерным участкам хромосом. Биваленты располагаются на периферии ядра. В конце профазы I ядерная оболочка разрушается, и биваленты выходят в цитоплазму.
Метафаза I: формируется веретено деления. Биваленты перемещаются в экваториальную плоскость клетки. Образуется метафазная пластинка из бивалентов.
Анафаза I: гомологичные хромосомы разъединяются, и каждая хромосома движется в сторону ближайшего полюса клетки. Разъединения хромосом на хроматиды не происходит.
Телофаза I: гомологичные двухроматидные хромосомы полностью расходятся к полюсам клетки. Формируются два гаплоидных ядра, которые содержат 2n2с.
Второе деление мейоза (эквационное или мейоз II)
Профаза II: не отличается существенно от профазы митоза. Хромосомы видны в световой микроскоп в виде тонких нитей. В каждой из дочерних клеток формируется веретено деления.
Метафаза II: хромосомы располагаются в экваториальных плоскостях.
Анафаза II: хромосомы разделяются на хроматиды (как при митозе). Получившиеся хроматиды перемещаются к полюсам клеток.
Телофаза II: однохроматидные хромосомы полностью переместились к полюсам клетки, формируются ядра. Содержание ДНК в каждой из клеток становится минимальным и составляет 1с.
Таким образом, в результате описанной схемы мейоза из одной диплоидной клетки образуется четыре гаплоидные клетки. Дальнейшая их судьба зависит от таксономической принадлежности организмов, от пола и других факторов.
Значение мейоза:
1. Поддержание постоянства числа хромосом. Если бы не возникала редукция числа хромосом при половом размножении, половые клетки не имели гаплоидный набор хромосом, то из поколения в поколение возрастало бы их число.
2. В процессе кроссинговера имеет место рекомбинация генетического материала. Практически все хромосомы, попадающие в гаметы, содержат участки, происходящие первоначально как от отцовской, так и от материнской хромосомы. Этим достигается большая степень перекомбинации наследственного материала. В этом одна из причин комбинативной изменчивости, дающей материал для отбора.
Отличия митоза от мейоза
При митозе в профазе конъюгация гомологичных хромосом бывает только в том случае, если гомологичные хромосомы окажутся рядом, и тогда может произойти кроссинговер. Удвоение хромосом соответствует каждому делению клетки в митозе.
В метафазе при митозе на экваторе выстраиваются хромосомы, состоящие из двух хроматид. В анафазе при митозе к полюсам расходятся хроматиды. В телофазе дочерние клетки содержат то же число хромосом, что и материнские.
При мейозе в профазе I происходит конъюгация гомологичных хромосом, имеет место кроссинговер. Образуются биваленты хромосом. В метафазе I при мейозе на экваторе располагаются биваленты хромосомы. При мейозе в анафазе I к полюсам расходятся хромосомы, состоящие из двух хроматид. В телофазе I мейоза число хромосом в дочерних клетках вдвое меньше, чем в материнских, редукционное деление.
Между I и II делениями мейоза в интерфазе нет синтеза ДНК. В результате мейоза при сперматогенезе образуются четыре клетки, при овогенезе одна половая клетка и три направительных (полярных) тельца, и все они будут иметь гаплоидный набор хромосом.
- Сперматогенез и овогенез. Особенности гаметогенеза у человека. Понятие о первичном и вторичном нерасхождении хромосом в гаметогенезе. Влияние пола, возраста, явления элиминации аномальных гамет на частоту нерасхождения хромосом в гаметогенезе.
Примордиальные зародышевые (герминативные) клетки, которые мигрируют в гонады во время раннего развития зародыша, размножаются путем митотического деления. Действительное образование половых клеток, гаметогенез, начинается с мейоза. Мейоз у самцов и самок отличается по длительности и результатам. Сходство сперматогенеза и овогенеза заключается в том, что три стадии у них одинаковы.
- Стадия размножения: сперматогонии и овогонии, несколько делений митоза, увеличение количества. Сперматогонии размножаются в течение всего репродуктивного периода, а размножение овогоний происходит в эмбриональном периоде и наиболее интенсивно происходит во 2-5 месяц развития.
- Стадия роста: сперматоциты и овоциты I порядка, увеличение в размерах. Овоциты I порядка достигают больших размеров, поскольку накапливают питательные вещества.
- Стадия созревания: на этой стадии происходят 2 мейоза. После первого деления образуются сперматоциты и овоциты II порядка, а после второго деления – сперматиды и зрелые яйцеклетки с тремя полярными тельцами. При созревании один сперматоцит I порядка дает четыре сперматиды, а один овоцит I порядка образует одну яйцеклетку и 2 полярных тельца.
- Стадия формирования: характерна только для сперматогенеза, сперматозоиды, начинается только в период полового созревания. Период развития составляет 74 дня.
Особенности гаметогенеза у человека:
1. Митотическое деление овогоний заканчивается до рождения организма. Митоз сперматогоний продолжается и в период полового созревания.
2. При овогенезе значительно выражена зона роста, при сперматогенезе зона роста почти не выражена.
3. При овогенезе первое деление мейоза останавливается на стадии диакинеза профазы до полового созревания. Второе деление мейоза останавливается на стадии метафазы и завершается после оплодотворения.
Родившаяся девочка имеет в яичниках около 30 000 ооцитов, достигают зрелости 300-600 (примерно по 13 клеток в год). За период половой жизни мужской организм продуцирует до 500 млрд сперматозоидов.
В настоящее время последние стадии овогенеза воспроизводятся вне организма и дают возможность «зачатия» в пробирке. На стадии 8-16 бластомеров зародыш переносится в матку женщины-реципиента.
У низших животных половые клетки вырабатываются в течение всей жизни, у высших – в период половой активности.
Основное преимущество полового размножения перед бесполым – увеличение генетического разнообразия видов и популяций.
Патология мейоза
Основная патология – нерасхождение хромосом. Оно может быть первичным, вторичным и третичным.
Первичное нерасхождение возникает у особей с нормальным кариотипом. При этом на стадии анафазы I нарушается разделение бивалентов и обе хромосомы из пары гомологов переходят в одну клетку, что приводит к избытку хромосом в данной клетке (n+1) и недостатку в другой (n-1).
Вторичное не расхождение возникает в гаметах у особей с избытком (трисомией) одной хромосомы в кариотипе. В результате этого в процессе мейоза образуются и биваленты, и униваленты.
Третичные нерасхождения наблюдают у особей, имеющих структурные перестройки хромосом, например транслокации. Нерасхождение хромосом отрицательно влияет на жизнеспособность организма животных.
Отличительная особенность мейоза у самок — образование в первом и втором мейотических делениях так называемых полярных телец, которые впоследствии дегенерируют и в размножении не участвуют. Неравные деления в овогенезе обеспечивают яйцеклетке необходимое количество цитоплазмы и запасного желтка, чтобы она могла выжить после оплодотворения. В отличие от спермиогенеза, который у самцов происходит как во внутриутробный (пренатальный) период, так и после рождения (постнатальный период), у самок яйцеклетка после рождения не образуется. Однако к концу пренатального периода у самок накапливается огромное количество овоцитов (у коров, например, десятки тысяч), а созревают и дают начало развитию потомства лишь немногие из них. Этот резерв генетического материала, особенно у малоплодных животных, таких, как крупный рогатый скот, в настоящее время начинают использовать для искусственного стимулирования созревания многих яйцеклеток, последующего их оплодотворения и пересадки (трансплантации) специально подготовленным коровам-реципиентам. Таким образом от одной ценной коровы в год можно получить не одного, а несколько телят.
- Морфология хромосом человека. Химический состав и молекулярная организация хромосом. Понятие об эухроматине. Понятие о нормальном хромосомном полиморфизме.
- Тератогенные факторы окружающей среды. Особенности тератогенного действия физических, химических и биологических факторов. Основные «ошибки» морфогенеза.
Тератогены – химические, физические и биологические факторы, способные нарушать процессы эмбриогенеза, что приводит к возникновению аномалий развития. Механизм возникновения пороков развития называется тератогенезом.
В возникновении уродства имеет значение как природа тератогенного фактора, так и видовые, индивидуальные, возрастные и другие особенности каждого организм. Один и тот же недостаток можно вызвать действием различных факторов и, наоборот, различные пороки возникают при действии одного и того же фактора.
Механические факторы: давление, сотрясения, механические травмы, различные виды радиации, особенно проникающей, гипо — и гипертермия. Ионизирующее излучение (один из самых тяжелых за эффектом тератогенов) в различных дозах вызывает различной степени нарушения развития и формы уродства. Ионизирующее излучение даже в малых дозах, не вызывая патологических сдвигов в организме, вызывает мутагенный эффект на половые клетки.
Химические тератогены:
- Нитраты, нитриты. Поступают в организм с переудобренными азотом овощами, в ЖКТ могут превращаться в нитриты. Нитриты, вступая в реакцию с аминами, образуют канцерогенные нитрозамины.
- Бензопирены. Образуются при жарке и приготовлении пищи на гриле. Их много в табачном дыме.
- Пероксиды. Образуются в прогорклых жирах и при сильном нагреве растительных масел.
- Афлатоксины. Продукты обмена плесневых грибов. Следует быть осторожным с заплесневелым хлебом и орехами и следить за влажностью в помещении.
- Диоксины. Хлорорганические соединения, образующиеся при сжигании бытового мусора. Образуются при хлорировании загрязнененной органикой воды (их можно получить из водопроводного крана весной, когда в поверхностные воды попадают талые воды с полей, удобренных навозом).
- Антибиотики (тетрациклин, оксациллин, антиметаболиты и др.), сульфаниламиды, варфарин и другие антикоагулянты, гормоны, противосудорожные, противоопухолевые и др.
- Спирты
- Растворители
- Витамины (гипер- и гиповитаминозы).
Особую заботу вызывают теперь употребление алкоголя, курение, злоупотребление наркотиками, которые негативно влияют на внутриутробное или постнатальное развитие.
Алкоголь – частый тератоген, а злоупотребление алкоголем во время беременности – частая причина химического индуцированного тератогенеза. Он непосредственная причина каждой 10-й эмбриональной патологии. Из 10 умственно неполноценных детей пятеро рождается от родителей-алкоголиков. Чрезвычайно широкий спектр аномалий проявляется у детей, рожденных матерями, которые злоупотребляли алкоголем: пороки развития сердца, почек, половых органов, кожи, скелета и суставов, анэнцефалии, гидроцефалии, микроцефалии, челюстно-лицевые аномалии.
Биологические тератогены: бактериальные токсины, вирусы (простой герпес, вирусный гепатит, грипп, краснуха, ветрянка, вирусы Коксаки, цитомегаловирус и др.), факторы иммунологической несовместимости, токсоплазмы.
Особенности влияния тератогенных факторов:
1. Действие тератогенных факторов имеет дозозависимый характер. У разных биологических видов дозозависимость тератогенного действия может различаться.
2. Для каждого тератогенного фактора существует определенная пороговая доза тератогенного действия. Обычно она на 1-3 порядка ниже летальной.
3. Различия тератогенного действия у различных биологических видов, а также у разных представителей одного и того же вида связаны с особенностями всасывания, метаболизма, способности вещества распространяться в организме и проникать через плаценту.
4. Чувствительность к разным тератогенным факторам в течение внутриутробного развития может меняться. Выделяют следующие периоды внутриутробного развития человека.
- оплодотворение;
- имплантацию (7-8-е сутки эмбриогенеза);
- развитие осевого комплекса зачатков органов и плацентацию (3-8-я недели);
- развитие головного мозга (15-20-я недели);
- формирование основных систем организма, в том числе половой (20-24-я недели);
5. В случаях, когда тератогенное действие оказывают возбудители инфекций, пороговую дозу и дозозависимый характер действия тератогенного фактора оценить не удается.
- Эпигенетическая наследственность. Импринтинг. Болезни, связанные с нарушением импринтинга.
Эпигенетическим наследованием называют наследуемые изменения в фенотипе или экспрессии генов, вызываемые механизмами, отличными от изменения последовательности ДНК.
В более узком смысле слово «эпигенетика» означает модификацию генной экспрессии, обусловленную наследственными, но потенциально обратимыми изменениями в структуре хроматина и/или в результате метилирования ДНК.
В случае эпигенетического наследования не происходит изменения последовательности ДНК, но другие генетические факторы регулируют активность генов. Лучшим примером эпигенетических изменений для эукариот является процесс дифференцировки клеток. В течение морфогенеза тотипотентные стволовые клетки становятся плюрипотентными линиями клеток, которые в тканях эмбриона затем превращаются в полностью дифференцированные клетки. В процессе дифференцировки активируются одни гены и инактивируются другие.
Эпигенетические изменения:
Процесс метилирования цитозина – наиболее изученный механизм эпигенетической регуляции активности генов, может влиять на активность генов несколькими способами. В частности, метильные группы могут физически препятствовать контакту фактора транскрипции со специфичными участками ДНК, могут участвовать в процессе ремоделирования хроматина. инактивации X-хромосом у эмбриона.
Образование конститутивного хроматина
Метилирование, фосфорилирование, ацетилирование гистонов
РНК-интерференция
Генетический импринтинг – механизм, с помощью которого различается активность генов в зависимости от того, от какого родителя они получены – материнского или отцовского организма. В участках генома, подверженных импринтингу, происходит моноаллельная экспрессия генов. В одних случаях импринтингу подвергаются отцовские гены, и, следовательно, транскрибируются материнские, а в других – материнские, что приводит к отклонению от менделевских законов. Речь идёт о стойких функциональных различиях экспрессии гомологичных генов у потомства.
Примеры:
- тяжелая (злокачественная) шизофрения – проявляется более выраженно, если наследуется по отцовской линии;
- нейрофиброматоз Реклингаузена (тип 2) – мутация в гене SCH (22q12); наследуется по материнской линии;
- псориаз – проявляется тяжелее, если наследуется по отцовской линии;
- синдром Прадера-Вилли (СПВ) – делеция критического района, находящегося в отцовской хромосоме 15 (15q11.2-q13) или синдром Ангельмана (СА) – делеция того же района, находящегося в материнской хромосоме;
- эпилепсия – проявляется тяжелее, если наследуется по материнской линии;
- синдром «крика кошки» — проявляется более выраженно, если делеция захватывает отцовскую хромосому 5.