

-

- 23 май 2016
- 11
- A4 2.0 TFSI Q 224 2014
Добрый вечер ребят, вот собираюсь купить Audi A4 2.0 TFSI Q 2014 г.в.
При проверке компьютером выдал такую ошибку:
Код неисправности: P11A600Описание: Исполнительный механизм переключения передач А, цилиндр 2: диапазон/качество работы — сигнал недостоверен.
Состояние: Пассивн/абиогенез
Кто что знает по данной ошибке? Авто на гарантии, но проводить большое кол-во времени у диллера просто физически не могу (Я из другого города где оф. диллеров нет)
-
Лучше обойди эту машину стороной
Stop hovering to collapse…
Click to collapse…
Hover to expand…
Нажмите, чтобы раскрыть…
-
- 19 июн 2010
- 19.986
- А6/С6 3.0i Q 05
Перевод дерьмовый, вот так правильно.
P11A600 — Camshaft position (CMP) actuator A, cylinder 2 — range/performance problem
К дилеру езжай, не осилишь без багажа опыта, даже не суйся.
Stop hovering to collapse…
Click to collapse…
Hover to expand…
Нажмите, чтобы раскрыть…
-
- 24 окт 2009
- 32.844
- A4B6 1.8TQ AT 04г
точно, покупай машину, и к дилеру сразу
-
- 23 май 2016
- 11
- A4 2.0 TFSI Q 224 2014
Я так и собирался делать, гарантии еще пол года.
Просто не пойму что это за проблема, вы не знаете? -
- 23 май 2016
- 11
- A4 2.0 TFSI Q 224 2014
-
- 24 окт 2009
- 32.844
- A4B6 1.8TQ AT 04г
чет такой ошибки гугель не знает..
-
- 23 май 2016
- 11
- A4 2.0 TFSI Q 224 2014
Да, искал. Нашел только один форум по VW Golf 2016 года, там такая же проблема, двигатель тот же. Но ему сказали что нужно просто самую последнюю прошивку поставить.
-
- 19 июн 2010
- 19.986
- А6/С6 3.0i Q 05
Знаю, но не вижу смысла впадать в дискуссию.
Stop hovering to collapse…
Click to collapse…
Hover to expand…
Нажмите, чтобы раскрыть…
-
- 24 окт 2009
- 32.844
- A4B6 1.8TQ AT 04г
Если в ЛНР, я бы там такую не брал, там и бенза ща то нормального нет наверное..
Тем более сервисов.. -
- 23 май 2016
- 11
- A4 2.0 TFSI Q 224 2014
-
- 23 май 2016
- 11
- A4 2.0 TFSI Q 224 2014
Ну сервисов да, бензин тоже думаю не самый лучший. Но в принципе, ездить раз в месяц в Ростовскую область думаю проблемой для меня не станет, так что страдать от бензина ЛНРовского не буду.
Машина 14 года, 17 тысяч пробега, верю в то что в ближайшее время она не будет капризничать ;D -
- 19 июн 2010
- 19.986
- А6/С6 3.0i Q 05
Скачай https://yadi.sk/d/gCVylKWVrzkry для понимания, а может и отремонтируешь сам.
Stop hovering to collapse…
Click to collapse…
Hover to expand…
Нажмите, чтобы раскрыть…
-
- 23 май 2016
- 11
- A4 2.0 TFSI Q 224 2014
Во, большое спасибо.
Сам делать не собирался, мне просто нужно было узнать на сколько данная проблема трудроемка (Долго ли будет чинить диллер)


«Origin of life» redirects here. For non-scientific views on the origins of life, see Creation myth.

Stages in the origin of life range from the well-understood, such as the habitable Earth and the abiotic synthesis of simple molecules, to the largely unknown, like the derivation of the last universal common ancestor (LUCA) with its complex molecular functionalities.[1]
In biology, abiogenesis (from a- ‘not’ + Greek bios ‘life’ + genesis ‘origin’) or the origin of life is the natural process by which life has arisen from non-living matter, such as simple organic compounds. The prevailing scientific hypothesis is that the transition from non-living to living entities on Earth was not a single event, but an evolutionary process (i.e., a process of gradually increasing complexity) that involved the formation of a habitable planet, the prebiotic synthesis of organic molecules, molecular self-replication, self-assembly, autocatalysis, and the emergence of cell membranes. Many proposals have been made for different stages of the process.
The study of abiogenesis aims to determine how pre-life chemical reactions gave rise to life under conditions strikingly different from those on Earth today. It primarily uses tools from biology and chemistry, with more recent approaches attempting a synthesis of many sciences. Life functions through the specialized chemistry of carbon and water, and builds largely upon four key families of chemicals: lipids for cell membranes, carbohydrates such as sugars, amino acids for protein metabolism, and nucleic acids DNA and RNA for the mechanisms of heredity. Any successful theory of abiogenesis must explain the origins and interactions of these classes of molecules. Many approaches to abiogenesis investigate how self-replicating molecules, or their components, came into existence. Researchers generally think that current life descends from an RNA world, although other self-replicating molecules may have preceded RNA.
The classic 1952 Miller–Urey experiment demonstrated that most amino acids, the chemical constituents of proteins, can be synthesized from inorganic compounds under conditions intended to replicate those of the early Earth. External sources of energy may have triggered these reactions, including lightning, radiation, atmospheric entries of micro-meteorites and implosion of bubbles in sea and ocean waves. Other approaches («metabolism-first» hypotheses) focus on understanding how catalysis in chemical systems on the early Earth might have provided the precursor molecules necessary for self-replication.
A genomics approach has sought to characterise the last universal common ancestor (LUCA) of modern organisms by identifying the genes shared by Archaea and Bacteria, members of the two major branches of life (where the Eukaryotes belong to the archaean branch in the two-domain system). 355 genes appear to be common to all life; their nature implies that the LUCA was anaerobic with the Wood–Ljungdahl pathway, deriving energy by chemiosmosis, and maintaining its hereditary material with DNA, the genetic code, and ribosomes. Although the LUCA lived over 4 billion years ago (4 Gya), researchers do not believe it was the first form of life. Earlier cells might have had a leaky membrane and been powered by a naturally-occurring proton gradient near a deep-sea white smoker hydrothermal vent.
Earth remains the only place in the universe known to harbor life, and fossil evidence from the Earth informs most studies of abiogenesis. The Earth was formed 4.54 Gya; the earliest undisputed evidence of life on Earth dates from at least 3.5 Gya. Fossil micro-organisms appear to have lived within hydrothermal vent precipitates dated 3.77 to 4.28 Gya from Quebec, soon after ocean formation 4.4 Gya during the Hadean.
Overview[edit]
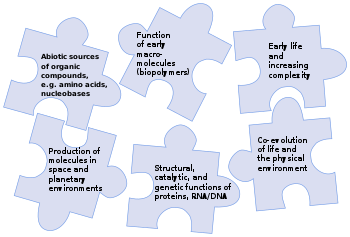
NASA’s 2015 strategy for astrobiology aimed to solve the puzzle of the origin of life – how a fully-functioning living system could emerge from non-living components – through research on the prebiotic origin of life’s chemicals, both in space and on planets, as well as the functioning of early biomolecules to catalyse reactions and support inheritance.[2]
Life consists of reproduction with (heritable) variations.[3] NASA defines life as «a self-sustaining chemical system capable of Darwinian [i.e., biological] evolution.»[4] Such a system is complex; the last universal common ancestor (LUCA), presumably a single-celled organism which lived some 4 billion years ago, already had hundreds of genes encoded in the DNA genetic code that is universal today. That in turn implies a suite of cellular machinery including messenger RNA, transfer RNAs, and ribosomes to translate the code into proteins. Those proteins included enzymes to operate its anaerobic respiration via the Wood–Ljungdahl metabolic pathway, and a DNA polymerase to replicate its genetic material.[5][6]
The challenge for abiogenesis (origin of life)[7][8][9] researchers is to explain how such a complex and tightly-interlinked system could develop by evolutionary steps, as at first sight all its parts are necessary to enable it to function. For example, a cell, whether the LUCA or in a modern organism, copies its DNA with the DNA polymerase enzyme, which is in turn produced by translating the DNA polymerase gene in the DNA. Neither the enzyme nor the DNA can be produced without the other.[10] The evolutionary process could have involved molecular self-replication, self-assembly such as of cell membranes, and autocatalysis.[5][6][11]
The precursors to the development of a living cell like the LUCA are clear enough, if disputed in their details: a habitable world is formed with a supply of minerals and liquid water. Prebiotic synthesis creates a range of simple organic compounds, which are assembled into polymers such as proteins and RNA. The process after the LUCA, too, is readily understood: biological evolution caused the development of a wide range of species with varied forms and biochemical capabilities. The derivation of living things such as the LUCA from simple components, however, is far from understood.[1]
Although Earth remains the only place where life is known,[12][13] the science of astrobiology seeks evidence of life on other planets. The 2015 NASA strategy on the origin of life aimed to solve the puzzle by identifying interactions, intermediary structures and functions, energy sources, and environmental factors that contributed to the diversity, selection, and replication of evolvable macromolecular systems,[2] and mapping the chemical landscape of potential primordial informational polymers. The advent of polymers that could replicate, store genetic information, and exhibit properties subject to selection was, it suggested, most likely a critical step in the emergence of prebiotic chemical evolution.[2] Those polymers derived, in turn, from simple organic compounds such as nucleobases, amino acids and sugars that could have been formed by reactions in the environment.[14][8][15][16] A successful theory of the origin of life must explain how all these chemicals came into being.[17]
Conceptual history until the 1960s[edit]
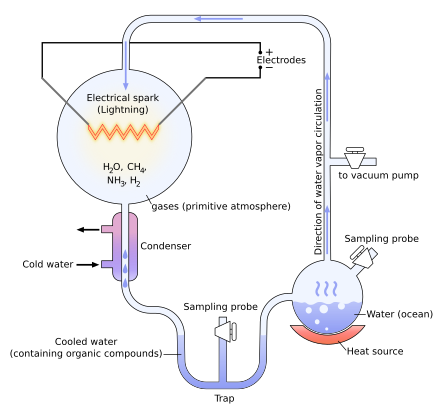
The Miller–Urey experiment was a synthesis of small organic molecules in a mixture of simple gases in a thermal gradient created by heating (right) and cooling (left) the mixture at the same time, with electrical discharges.
Spontaneous generation[edit]
One ancient view of the origin of life, from Aristotle until the 19th century, is of spontaneous generation.[18] This theory held that «lower» animals were generated by decaying organic substances, and that life arose by chance.[19][20] This was questioned from the 17th century, in works like Thomas Browne’s Pseudodoxia Epidemica.[21][22] In 1665, Robert Hooke published the first drawings of a microorganism. In 1676, Antonie van Leeuwenhoek drew and described microorganisms, probably protozoa and bacteria.[23] Van Leeuwenhoek disagreed with spontaneous generation, and by the 1680s convinced himself, using experiments ranging from sealed and open meat incubation and the close study of insect reproduction, that the theory was incorrect.[24] In 1668 Francesco Redi showed that no maggots appeared in meat when flies were prevented from laying eggs.[25] By the middle of the 19th century, spontaneous generation was considered disproven.[26][27]
Panspermia[edit]
Another ancient idea dating back to Anaxagoras in the 5th century BC is panspermia,[28] the idea that life exists throughout the universe, distributed by meteoroids, asteroids, comets[29] and planetoids.[30] It does not attempt to explain how life originated in itself, but shifts the origin of life on Earth to another heavenly body. The advantage is that life is not required to have formed on each planet it occurs on, but rather in a more limited set of locations (potentially even a single location), and then spread about the galaxy to other star systems via cometary or meteorite impact.[31]
«A warm little pond»: primordial soup[edit]
The idea that life originated from non-living matter in slow stages appeared in Herbert Spencer’s 1864–1867 book Principles of Biology, and in William Turner Thiselton-Dyer’s 1879 paper «On spontaneous generation and evolution». On 1 February 1871 Charles Darwin wrote about these publications to Joseph Hooker, and set out his own speculation, suggesting that the original spark of life may have begun in a «warm little pond, with all sorts of ammonia and phosphoric salts, light, heat, electricity, &c., present, that a proteine compound was chemically formed ready to undergo still more complex changes.» Darwin went on to explain that «at the present day such matter would be instantly devoured or absorbed, which would not have been the case before living creatures were formed.»[32][33][34]
Alexander Oparin in 1924 and J. B. S. Haldane in 1929 proposed that the first molecules constituting the earliest cells slowly self-organized from a primordial soup, and this theory is called the Oparin–Haldane hypothesis.[35][36] Haldane suggested that the Earth’s prebiotic oceans consisted of a «hot dilute soup» in which organic compounds could have formed.[20][37] J. D. Bernal showed that such mechanisms could form most of the necessary molecules for life from inorganic precursors.[38] In 1967, he suggested three «stages»: the origin of biological monomers; the origin of biological polymers; and the evolution from molecules to cells.[39][40]
Miller–Urey experiment[edit]
In 1952, Stanley Miller and Harold Urey carried out a chemical experiment to demonstrate how organic molecules could have formed spontaneously from inorganic precursors under prebiotic conditions like those posited by the Oparin–Haldane hypothesis. It used a highly reducing (lacking oxygen) mixture of gases—methane, ammonia, and hydrogen, as well as water vapor—to form simple organic monomers such as amino acids.[41][42] Bernal said of the Miller–Urey experiment that «it is not enough to explain the formation of such molecules, what is necessary, is a physical-chemical explanation of the origins of these molecules that suggests the presence of suitable sources and sinks for free energy.»[43] However, current scientific consensus describes the primitive atmosphere as weakly reducing or neutral,[44][45] diminishing the amount and variety of amino acids that could be produced. The addition of iron and carbonate minerals, present in early oceans, however produces a diverse array of amino acids.[44] Later work has focused on two other potential reducing environments: outer space and deep-sea hydrothermal vents.[46][47][48]
Producing a habitable Earth[edit]
Early universe with first stars[edit]
Soon after the Big Bang, which occurred roughly 14 Gya, the only chemical elements present in the universe were hydrogen, helium, and lithium, the three lightest atoms in the periodic table. These elements gradually came together to form stars. These early stars were massive and short-lived, producing all the heavier elements through stellar nucleosynthesis. Carbon, currently the fourth most abundant chemical element in the universe (after hydrogen, helium and oxygen), was formed mainly in white dwarf stars, particularly those bigger than twice the mass of the sun.[49] As these stars reached the end of their lifecycles, they ejected these heavier elements, among them carbon and oxygen, throughout the universe. These heavier elements allowed for the formation of new objects, including rocky planets and other bodies.[50] According to the nebular hypothesis, the formation and evolution of the Solar System began 4.6 Gya with the gravitational collapse of a small part of a giant molecular cloud. Most of the collapsing mass collected in the center, forming the Sun, while the rest flattened into a protoplanetary disk out of which the planets, moons, asteroids, and other small Solar System bodies formed.[51]
Emergence of Earth[edit]
The Earth was formed 4.54 Gya.[52][53] The Hadean Earth (from its formation until 4 Gya) was at first inhospitable to any living organisms. During its formation, the Earth lost a significant part of its initial mass, and consequentially lacked the gravity to hold molecular hydrogen and the bulk of the original inert gases.[54] The atmosphere consisted largely of water vapor, nitrogen, and carbon dioxide, with smaller amounts of carbon monoxide, hydrogen, and sulfur compounds.[55] The solution of carbon dioxide in water is thought to have made the seas slightly acidic, with a pH of about 5.5.[56] The Hadean atmosphere has been characterized as a «gigantic, productive outdoor chemical laboratory,»[57] similar to volcanic gases today which still support some abiotic chemistry.[57]
Oceans may have appeared as soon as 200 million years after the Earth formed, in a near-boiling (100 C) reducing environment, as the pH of 5.8 rose rapidly towards neutral.[58] This scenario has found support from the dating of 4.404 Gya zircon crystals from metamorphosed quartzite of Mount Narryer in Western Australia.[59] Despite the likely increased volcanism, the Earth may have been a water world between 4.4 and 4.3 Gya, with little if any continental crust, a turbulent atmosphere, and a hydrosphere subject to intense ultraviolet light from a T Tauri stage Sun, from cosmic radiation, and from continued asteroid and comet impacts.[60]
The Late Heavy Bombardment hypothesis posits that the Hadean environment between 4.28[61] and 3.8 Gya was highly hazardous to life. Following the Nice model, changes in the orbits of the giant planets may have bombarded the Earth with asteroids and comets that pockmarked the Moon and inner planets.[62] Frequent collisions would have made photosynthesis unviable.[57][63][64][65] The periods between such devastating events give time windows for the possible origin of life in early environments. If the deep marine hydrothermal setting was the site for the origin of life, then abiogenesis could have happened as early as 4.0-4.2 Gya. If the site was at the surface of the Earth, abiogenesis could have occurred only between 3.7 and 4.0 Gya.[66] However, new lunar surveys and samples have led scientists, including an architect of the Nice model, to deemphasize the significance of the Late Heavy Bombardment.[67]
If life evolved in the ocean at depths of more than ten meters, it would have been shielded both from late impacts and the then high levels of ultraviolet radiation from the sun. Geothermically heated oceanic crust could have yielded far more organic compounds through deep hydrothermal vents than the Miller–Urey experiments indicated.[68] The available energy is maximized at 100–150 °C, the temperatures at which hyperthermophilic bacteria and thermoacidophilic archaea live. These modern organisms may be among the closest surviving relatives of the LUCA.[69]
Earliest evidence of life[edit]
Life existed on Earth more than 3.5 Gya,[70][71][72] during the Eoarchean when sufficient crust had solidified following the molten Hadean.[73][74][75] The earliest physical evidence of life so far found consists of microfossils in the Nuvvuagittuq Greenstone Belt of Northern Quebec, in banded iron formation rocks at least 3.77 and possibly 4.28 Gya. The micro-organisms lived within hydrothermal vent precipitates, soon after the 4.4 Gya formation of oceans during the Hadean. The microbes resembled modern hydrothermal vent bacteria, supporting the view that abiogenesis began in such an environment.[61]
Biogenic graphite has been found in 3.7 Gya metasedimentary rocks from southwestern Greenland[76] and in microbial mat fossils from 3.49 Gya Western Australian sandstone.[77] Evidence of early life in rocks from Akilia Island, near the Isua supracrustal belt in southwestern Greenland, dating to 3.7 Gya, have shown biogenic carbon isotopes.[78] In other parts of the Isua supracrustal belt, graphite inclusions trapped within garnet crystals are connected to the other elements of life: oxygen, nitrogen, and possibly phosphorus in the form of phosphate, providing further evidence for life 3.7 Gya.[79] In the Pilbara region of Western Australia, compelling evidence of early life was found in pyrite-bearing sandstone in a fossilized beach, with rounded tubular cells that oxidized sulfur by photosynthesis in the absence of oxygen.[80][81] Zircons from Western Australia imply that life existed on Earth at least 4.1 Gya.[82]
The Pilbara region of Western Australia contains the Dresser Formation with rocks 3.48 Gya, including layered structures called stromatolites. Their modern counterparts are created by photosynthetic micro-organisms including cyanobacteria.[83] These lie within undeformed hydrothermal-sedimentary strata; their texture indicates a biogenic origin. Parts of the Dresser formation preserve hot springs on land, but other regions seem to have been shallow seas.[84]
Producing molecules: prebiotic synthesis[edit]
All chemical elements except for hydrogen and helium derive from stellar nucleosynthesis. The basic chemical ingredients of life – the carbon-hydrogen molecule (CH), the carbon-hydrogen positive ion (CH+) and the carbon ion (C+) – were produced by ultraviolet light from stars.[85] Complex molecules, including organic molecules, form naturally both in space and on planets.[86] Organic molecules on the early Earth could have had either terrestrial origins, with organic molecule synthesis driven by impact shocks or by other energy sources, such as ultraviolet light, redox coupling, or electrical discharges; or extraterrestrial origins (pseudo-panspermia), with organic molecules formed in interstellar dust clouds raining down on to the planet.[87][88]
[edit]
An organic compound is a chemical whose molecules contain carbon. Carbon is abundant in the Sun, stars, comets, and in the atmospheres of most planets.[89] Organic compounds are relatively common in space, formed by «factories of complex molecular synthesis» which occur in molecular clouds and circumstellar envelopes, and chemically evolve after reactions are initiated mostly by ionizing radiation.[86][90][91] Purine and pyrimidine nucleobases including guanine, adenine, cytosine, uracil and thymine have been found in meteorites. These could have provided the materials for DNA and RNA to form on the early Earth.[92] The amino acid glycine was found in material ejected from comet Wild 2; it had earlier been detected in meteorites.[93] Comets are encrusted with dark material, thought to be a tar-like organic substance formed from simple carbon compounds under ionizing radiation. A rain of material from comets could have brought such complex organic molecules to Earth.[94][95][57] It is estimated that during the Late Heavy Bombardment, meteorites may have delivered up to five million tons of organic prebiotic elements to Earth per year.[57]
PAH world hypothesis[edit]

Polycyclic aromatic hydrocarbons (PAH) are the most common and abundant polyatomic molecules in the observable universe, and are a major store of carbon.[89][96][97][98] They seem to have formed shortly after the Big Bang,[99][97][98] and are associated with new stars and exoplanets.[89] They are a likely constituent of Earth’s primordial sea.[99][97][98] PAHs have been detected in nebulae,[100] and in the interstellar medium, in comets, and in meteorites.[89]
The PAH world hypothesis posits PAHs as precursors to the RNA world.[101] A star, HH 46-IR, resembling the sun early in its life, is surrounded by a disk of material which contains molecules including cyanide compounds, hydrocarbons, and carbon monoxide. PAHs in the interstellar medium can be transformed through hydrogenation, oxygenation, and hydroxylation to more complex organic compounds used in living cells.[102]
Nucleobases[edit]
The majority of organic compounds introduced on Earth by interstellar dust particles have helped to form complex molecules, thanks to their peculiar surface-catalytic activities.[103][104] Studies of the 12C/13C isotopic ratios of organic compounds in the Murchison meteorite suggest that the RNA component uracil and related molecules, including xanthine, were formed extraterrestrially.[105] NASA studies of meteorites suggest that all four DNA nucleobases (adenine, guanine and related organic molecules) have been formed in outer space.[103][106][107] The cosmic dust permeating the universe contains complex organics («amorphous organic solids with a mixed aromatic–aliphatic structure») that could be created rapidly by stars.[108] Glycolaldehyde, a sugar molecule and RNA precursor, has been detected in regions of space including around protostars and on meteorites.[109][110]
Laboratory synthesis[edit]
As early as the 1860s, experiments demonstrated that biologically relevant molecules can be produced from interaction of simple carbon sources with abundant inorganic catalysts. The spontaneous formation of complex polymers from abiotically generated monomers under the conditions posited by the «soup» theory is not straightforward. Besides the necessary basic organic monomers, compounds that would have prohibited the formation of polymers were also formed in high concentration during the Miller–Urey and Joan Oró experiments.[111] Biology uses essentially 20 amino acids for its coded protein enzymes, representing a very small subset of the structurally possible products. Since life tends to use whatever is available, an explanation is needed for why the set used is so small.[112]
Sugars[edit]

The Breslow catalytic cycle for formaldehyde dimerization and C2-C6 sugar formation
Alexander Butlerov showed in 1861 that the formose reaction created sugars including tetroses, pentoses, and hexoses when formaldehyde is heated under basic conditions with divalent metal ions like calcium. R. Breslow proposed that the reaction was autocatalytic in 1959.[113]
Nucleobases[edit]
Nucleobases like guanine and adenine can be synthesized from simple carbon and nitrogen sources like hydrogen cyanide (HCN) and ammonia.[114] Formamide produces all four ribonucleotides when warmed with terrestrial minerals. Formamide is ubiquitous in the Universe, produced by the reaction of water and HCN. It can be concentrated by the evaporation of water.[115][116] HCN is poisonous only to aerobic organisms (eukaryotes and aerobic bacteria), which did not yet exist. It can play roles in other chemical processes such as the synthesis of the amino acid glycine.[57]
DNA and RNA components including uracil, cytosine and thymine can be synthesized under outer space conditions, using starting chemicals such as pyrimidine found in meteorites. Pyrimidine may have been formed in red giant stars or in interstellar dust and gas clouds.[117] All four RNA-bases may be synthesized from formamide in high-energy density events like extraterrestrial impacts.[118]
Other pathways for synthesizing bases from inorganic materials have been reported.[119] Freezing temperatures are advantageous for the synthesis of purines, due to the concentrating effect for key precursors such as hydrogen cyanide.[120] However, while adenine and guanine require freezing conditions for synthesis, cytosine and uracil may require boiling temperatures.[121] Seven different amino acids and eleven types of nucleobases formed in ice when ammonia and cyanide were left in a freezer for 25 years.[122][123] S-triazines (alternative nucleobases), pyrimidines including cytosine and uracil, and adenine can be synthesized by subjecting a urea solution to freeze-thaw cycles under a reductive atmosphere, with spark discharges as an energy source.[124] The explanation given for the unusual speed of these reactions at such a low temperature is eutectic freezing, which crowds impurities in microscopic pockets of liquid within the ice, causing the molecules to collide more often.[125]
Producing suitable vesicles[edit]

The lipid world theory postulates that the first self-replicating object was lipid-like.[126][127] Phospholipids form lipid bilayers in water while under agitation—the same structure as in cell membranes. These molecules were not present on early Earth, but other amphiphilic long-chain molecules also form membranes. These bodies may expand by insertion of additional lipids, and may spontaneously split into two offspring of similar size and composition. The main idea is that the molecular composition of the lipid bodies is a preliminary to information storage, and that evolution led to the appearance of polymers like RNA that store information. Studies on vesicles from amphiphiles that might have existed in the prebiotic world have so far been limited to systems of one or two types of amphiphiles.[128]
A lipid bilayer membrane could be composed of a huge number of combinations of arrangements of amphiphiles. The best of these would have favored the constitution of a hypercycle,[129][130] actually a positive feedback composed of two mutual catalysts represented by a membrane site and a specific compound trapped in the vesicle. Such site/compound pairs are transmissible to the daughter vesicles leading to the emergence of distinct lineages of vesicles, which would have allowed natural selection.[131]
A protocell is a self-organized, self-ordered, spherical collection of lipids proposed as a stepping-stone to the origin of life.[128] The theory of classical irreversible thermodynamics treats self-assembly under a generalized chemical potential within the framework of dissipative systems.[132][133][134]
A central question in evolution is how simple protocells first arose and differed in reproductive contribution to the following generation, thus driving the evolution of life. A functional protocell has (as of 2014) not yet been achieved in a laboratory setting.[135][136][137] Self-assembled vesicles are essential components of primitive cells.[128] The second law of thermodynamics requires that the universe move in a direction in which entropy increases, yet life is distinguished by its great degree of organization. Therefore, a boundary is needed to separate life processes from non-living matter.[138] Irene Chen and Jack W. Szostak suggest that elementary protocells can give rise to cellular behaviors including primitive forms of differential reproduction, competition, and energy storage.[136] Competition for membrane molecules would favor stabilized membranes, suggesting a selective advantage for the evolution of cross-linked fatty acids and even the phospholipids of today.[136] Such micro-encapsulation would allow for metabolism within the membrane and the exchange of small molecules, while retaining large biomolecules inside. Such a membrane is needed for a cell to create its own electrochemical gradient to store energy by pumping ions across the membrane.[139][140]
Producing biology[edit]
Energy and entropy[edit]
Life requires a loss of entropy, or disorder, when molecules organize themselves into living matter. The emergence of life and increased complexity does not contradict the second law of thermodynamics, which states that overall entropy never decreases, since a living organism creates order in some places (e.g. its living body) at the expense of an increase of entropy elsewhere (e.g. heat and waste production).[141][142][143]
Multiple sources of energy were available for chemical reactions on the early Earth. Heat from geothermal processes is a standard energy source for chemistry. Other examples include sunlight, lightning,[57] atmospheric entries of micro-meteorites,[144] and implosion of bubbles in sea and ocean waves.[145] This has been confirmed by experiments[146][147] and simulations.[148]
Unfavorable reactions can be driven by highly favorable ones, as in the case of iron-sulfur chemistry. For example, this was probably important for carbon fixation.[a] Carbon fixation by reaction of CO2 with H2S via iron-sulfur chemistry is favorable, and occurs at neutral pH and 100 °C. Iron-sulfur surfaces, which are abundant near hydrothermal vents, can drive the production of small amounts of amino acids and other biomolecules.[57]
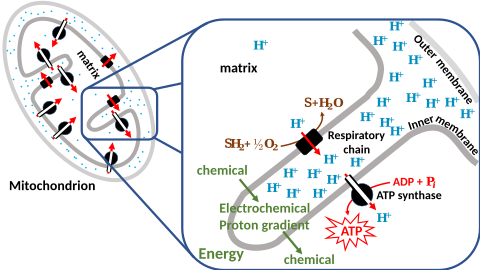
Chemiosmosis[edit]

In 1961, Peter Mitchell proposed chemiosmosis as a cell’s primary system of energy conversion. The mechanism, now ubiquitous in living cells, powers energy conversion in micro-organisms and in the mitochondria of eukaryotes, making it a likely candidate for early life.[149][150] Mitochondria produce adenosine triphosphate (ATP), the energy currency of the cell used to drive cellular processes such as chemical syntheses. The mechanism of ATP synthesis involves a closed membrane in which the ATP synthase enzyme is embedded. The energy required to release strongly-bound ATP has its origin in protons that move across the membrane.[151] In modern cells, those proton movements are caused by the pumping of ions across the membrane, maintaining an electrochemical gradient. In the first organisms, the gradient could have been provided by the difference in chemical composition between the flow from a hydrothermal vent and the surrounding seawater,[140] or perhaps meteoric quinones that were conducive to the development of chemiosmotic energy across lipid membranes if at a terrestrial origin.[152]
The RNA world[edit]
The RNA world hypothesis describes an early Earth with self-replicating and catalytic RNA but no DNA or proteins.[153] Many researchers concur that an RNA world must have preceded the DNA-based life that now dominates.[154] However, RNA-based life may not have been the first to exist.[155][156] Another model echoes Darwin’s «warm little pond» with cycles of wetting and drying.[157]
RNA is central to the translation process. Small RNAs can catalyze all the chemical groups and information transfers required for life.[156][158] RNA both expresses and maintains genetic information in modern organisms; and the chemical components of RNA are easily synthesized under the conditions that approximated the early Earth, which were very different from those that prevail today. The structure of the ribozyme has been called the «smoking gun», with a central core of RNA and no amino acid side chains within 18 Å of the active site that catalyzes peptide bond formation.[159][155][160]
The concept of the RNA world was proposed in 1962 by Alexander Rich,[161] and the term was coined by Walter Gilbert in 1986.[156][162] There were initial difficulties in the explanation of the abiotic synthesis of the nucleotides cytosine and uracil.[163] Subsequent research has shown possible routes of synthesis; for example, formamide produces all four ribonucleotides and other biological molecules when warmed in the presence of various terrestrial minerals.[115][116]
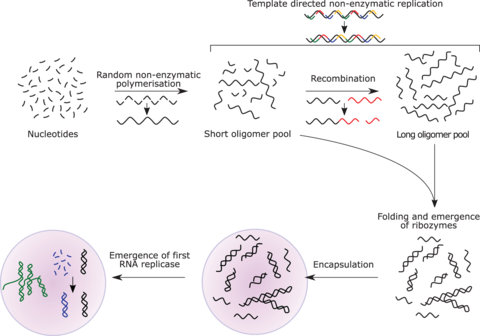
The RNA world hypothesis proposes that undirected polymerisation led to the emergence of ribozymes, and in turn to an RNA replicase.
RNA replicase can function as both code and catalyst for further RNA replication, i.e. it can be autocatalytic. Jack Szostak has shown that certain catalytic RNAs can join smaller RNA sequences together, creating the potential for self-replication. The RNA replication systems, which include two ribozymes that catalyze each other’s synthesis, showed a doubling time of the product of about one hour, and were subject to natural selection under the experimental conditions.[164][165][155] If such conditions were present on early Earth, then natural selection would favor the proliferation of such autocatalytic sets, to which further functionalities could be added.[166][167][168] Self-assembly of RNA may occur spontaneously in hydrothermal vents.[169][170][171] A preliminary form of tRNA could have assembled into such a replicator molecule.[172]
Possible precursors to protein synthesis include the synthesis of short peptide cofactors or the self-catalysing duplication of RNA. It is likely that the ancestral ribosome was composed entirely of RNA, although some roles have since been taken over by proteins. Major remaining questions on this topic include identifying the selective force for the evolution of the ribosome and determining how the genetic code arose.[173]
Eugene Koonin has argued that «no compelling scenarios currently exist for the origin of replication and translation, the key processes that together comprise the core of biological systems and the apparent pre-requisite of biological evolution. The RNA World concept might offer the best chance for the resolution of this conundrum but so far cannot adequately account for the emergence of an efficient RNA replicase or the translation system.»[174]
Phylogeny and LUCA[edit]
Starting with the work of Carl Woese from 1977 onwards, genomics studies have placed the last universal common ancestor (LUCA) of all modern life-forms between Bacteria and a clade formed by Archaea and Eukaryota in the phylogenetic tree of life. It lived over 4 Gya.[175][176] A minority of studies have placed the LUCA in Bacteria, proposing that Archaea and Eukaryota are evolutionarily derived from within Eubacteria;[177] Thomas Cavalier-Smith suggested that the phenotypically diverse bacterial phylum Chloroflexota contained the LUCA.[178]
In 2016, a set of 355 genes likely present in the LUCA was identified. A total of 6.1 million prokaryotic genes from Bacteria and Archaea were sequenced, identifying 355 protein clusters from amongst 286,514 protein clusters that were probably common to the LUCA. The results suggest that the LUCA was anaerobic with a Wood–Ljungdahl pathway, nitrogen- and carbon-fixing, thermophilic. Its cofactors suggest dependence upon an environment rich in hydrogen, carbon dioxide, iron, and transition metals. Its genetic material was probably DNA, requiring the 4-nucleotide genetic code, messenger RNA, transfer RNAs, and ribosomes to translate the code into proteins such as enzymes. LUCA likely inhabited an anaerobic hydrothermal vent setting in a geochemically active environment. It was evidently already a complex organism, and must have had precursors; it was not the first living thing.[10][179] The physiology of LUCA has been in dispute.[180][181][182]
-
![LUCA systems and environment[10]](data:image/svg+xml,%3Csvg%20xmlns='http://www.w3.org/2000/svg'%20viewBox='0%200%20415%20300'%3E%3C/svg%3E)
LUCA systems and environment[10]
![LUCA systems and environment[10]](https://upload.wikimedia.org/wikipedia/commons/thumb/e/ed/LUCA_systems_and_environment.svg/622px-LUCA_systems_and_environment.svg.png)
Leslie Orgel argued that early translation machinery for the genetic code would be susceptible to error catastrophe. Geoffrey Hoffmann however showed that such machinery can be stable in function against «Orgel’s paradox».[183][184][185]
Suitable geological environments[edit]
Deep sea hydrothermal vents[edit]

Early micro-fossils may have come from a hot world of gases such as methane, ammonia, carbon dioxide and hydrogen sulphide, toxic to much current life.[186] Analysis of the tree of life places thermophilic and hyperthermophilic bacteria and archaea closest to the root, suggesting that life may have evolved in a hot environment.[187] The deep sea or alkaline hydrothermal vent theory posits that life began at submarine hydrothermal vents.[188][189] Martin and Russell have suggested «that life evolved in structured iron monosulphide precipitates in a seepage site hydrothermal mound at a redox, pH, and temperature gradient between sulphide-rich hydrothermal fluid and iron(II)-containing waters of the Hadean ocean floor. The naturally arising, three-dimensional compartmentation observed within fossilized seepage-site metal sulphide precipitates indicates that these inorganic compartments were the precursors of cell walls and membranes found in free-living prokaryotes. The known capability of FeS and NiS to catalyze the synthesis of the acetyl-methylsulphide from carbon monoxide and methylsulphide, constituents of hydrothermal fluid, indicates that pre-biotic syntheses occurred at the inner surfaces of these metal-sulphide-walled compartments».[190]
These form where hydrogen-rich fluids emerge from below the sea floor, as a result of serpentinization of ultra-mafic olivine with seawater and a pH interface with carbon dioxide-rich ocean water. The vents form a sustained chemical energy source derived from redox reactions, in which electron donors (molecular hydrogen) react with electron acceptors (carbon dioxide); see Iron–sulfur world theory. These are exothermic reactions.[188][b]
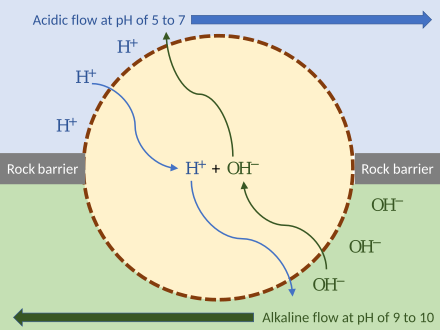
Early cell powered by external proton gradient near a deep-sea hydrothermal vent. As long as the membrane (or passive ion channels within it) is permeable to protons, the mechanism can function without ion pumps.[140]
Russell demonstrated that alkaline vents created an abiogenic proton motive force chemiosmotic gradient,[190] ideal for abiogenesis. Their microscopic compartments «provide a natural means of concentrating organic molecules,» composed of iron-sulfur minerals such as mackinawite, endowed these mineral cells with the catalytic properties envisaged by Günter Wächtershäuser.[191] This movement of ions across the membrane depends on a combination of two factors:
- Diffusion force caused by concentration gradient—all particles including ions tend to diffuse from higher concentration to lower.
- Electrostatic force caused by electrical potential gradient—cations like protons H+ tend to diffuse down the electrical potential, anions in the opposite direction.
These two gradients taken together can be expressed as an electrochemical gradient, providing energy for abiogenic synthesis. The proton motive force can be described as the measure of the potential energy stored as a combination of proton and voltage gradients across a membrane (differences in proton concentration and electrical potential).[140]
The surfaces of mineral particles inside deep-ocean hydrothermal vents have catalytic properties similar to those of enzymes and can create simple organic molecules, such as methanol (CH3OH), formic, acetic and pyruvic acids out of the dissolved CO2 in the water, if driven by an applied voltage or by reaction with H2 or H2S.[192][193]
The research reported by Martin in 2016 supports the thesis that life arose at hydrothermal vents,[194][195] that spontaneous chemistry in the Earth’s crust driven by rock–water interactions at disequilibrium thermodynamically underpinned life’s origin[196][197] and that the founding lineages of the archaea and bacteria were H2-dependent autotrophs that used CO2 as their terminal acceptor in energy metabolism.[198] Martin suggests, based upon this evidence, that the LUCA «may have depended heavily on the geothermal energy of the vent to survive».[199] Pores at deep sea hydrothermal vents are suggested to have been occupied
by membrane-bound compartments which promoted biochemical reactions.[200][201]
Hot springs[edit]
Mulkidjanian and co-authors think that marine environments did not provide the ionic balance and composition universally found in cells, or the ions required by essential proteins and ribozymes, especially with respect to high K+/Na+ ratio, Mn2+, Zn2+ and phosphate concentrations. They argue that the only environments that mimic the needed conditions on Earth are hot springs similar to ones at Kamchatka.[202] Mineral deposits in these environments under an anoxic atmosphere would have suitable pH (while current pools in an oxygenated atmosphere would not), contain precipitates of photocatalytic sulfide minerals that absorb harmful ultraviolet radiation, have wet-dry cycles that concentrate substrate solutions to concentrations amenable to spontaneous formation of biopolymers[203][204] created both by chemical reactions in the hydrothermal environment, and by exposure to UV light during transport from vents to adjacent pools that would promote the formation of biomolecules.[205] The hypothesized pre-biotic environments are similar to hydrothermal vents, with additional components that help explain peculiarities of the LUCA.[202][152]
A phylogenomic and geochemical analysis of proteins plausibly traced to the LUCA shows that the ionic composition of its intracellular fluid is identical at hot springs. The LUCA likely was dependent upon synthesized organic matter for its growth.[202] Experiments show that RNA-like polymers can be synthesized in multiple wet-dry cycles and exposure to UV light. These polymers became encapsulated in vesicles after condensation, which would not happen in saltwater conditions because of the high concentrations of ionic solutes.[206] A potential source of biomolecules at hot springs was transport by interplanetary dust particles, extraterrestrial projectiles, or atmospheric or geochemical synthesis. Hot spring fields could have been abundant at volcanic landmasses during the Hadean.[152]
Clay[edit]
The clay hypothesis was proposed by Graham Cairns-Smith in 1985.[207][208] It postulates that complex organic molecules arose gradually on pre-existing, non-organic replication surfaces of silicate crystals in contact with an aqueous solution. The clay mineral montmorillonite has been shown to catalyze the polymerization of RNA in aqueous solution from nucleotide monomers,[209] and the formation of membranes from lipids.[210] In 1998, Hyman Hartman proposed that «the first organisms were self-replicating iron-rich clays which fixed carbon dioxide into oxalic acid and other dicarboxylic acids. This system of replicating clays and their metabolic phenotype then evolved into the sulfide rich region of the hot spring acquiring the ability to fix nitrogen. Finally phosphate was incorporated into the evolving system which allowed the synthesis of nucleotides and phospholipids.»[211]
Iron–sulfur world[edit]
In the 1980s, Günter Wächtershäuser and Karl Popper postulated the Iron–sulfur world hypothesis for the evolution of pre-biotic chemical pathways. It traces today’s biochemistry to primordial reactions which synthesize organic building blocks from gases.[212][213] Wächtershäuser systems have a built-in source of energy: iron sulfides such as pyrite. The energy released by oxidising these metal sulfides can support synthesis of organic molecules. Such systems may have evolved into autocatalytic sets constituting self-replicating, metabolically active entities predating modern life forms.[214] Experiments with sulfides in an aqueous environment at 100 °C produced a small yield of dipeptides (0.4% to 12.4%) and a smaller yield of tripeptides (0.10%). However, under the same conditions, dipeptides were quickly broken down.[215]
Several models postulate a primitive metabolism, allowing RNA replication to emerge later. The centrality of the Krebs cycle (citric acid cycle) to energy production in aerobic organisms, and in drawing in carbon dioxide and hydrogen ions in biosynthesis of complex organic chemicals, suggests that it was one of the first parts of the metabolism to evolve.[191] Concordantly, geochemists Jack W. Szostak and Kate Adamala demonstrated that non-enzymatic RNA replication in primitive protocells is only possible in the presence of weak cation chelators like citric acid. This provides further evidence for the central role of citric acid in primordial metabolism.[216]
Russell has proposed that «the purpose of life is to hydrogenate carbon dioxide» (as part of a «metabolism-first,» rather than a «genetics-first,» scenario).[217][218][214] The physicist Jeremy England has argued from general thermodynamic considerations that life was inevitable.[219] An early version of this idea was Oparin’s 1924 proposal for self-replicating vesicles. In the 1980s and 1990s came Wächtershäuser’s iron–sulfur world theory and Christian de Duve’s thioester models. More abstract and theoretical arguments for metabolism without genes include Freeman Dyson’s mathematical model and Stuart Kauffman’s collectively autocatalytic sets in the 1980s. Kauffman’s work has been criticized for ignoring the role of energy in driving biochemical reactions in cells.[220]
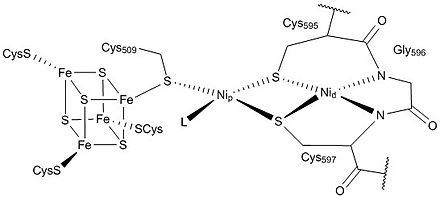
A multistep biochemical pathway like the Krebs cycle did not just self-organize on the surface of a mineral; it must have been preceded by simpler pathways.[221] The Wood–Ljungdahl pathway is compatible with self-organization on a metal sulfide surface. Its key enzyme unit, carbon monoxide dehydrogenase/acetyl-CoA synthase, contains mixed nickel-iron-sulfur clusters in its reaction centers and catalyzes the formation of acetyl-CoA. However, prebiotic thiolated and thioester compounds are thermodynamically and kinetically unlikely to accumulate in the presumed prebiotic conditions of hydrothermal vents.[222] One possibility is that cysteine and homocysteine may have reacted with nitriles from the Stecker reaction, forming catalytic thiol-rich polypeptides.[223]
Zinc world[edit]
Armen Mulkidjanian’s zinc world (Zn-world) hypothesis extends Wächtershäuser’s pyrite hypothesis.[224][225] The Zn-world theory proposes that hydrothermal fluids rich in H2S interacting with cold primordial ocean (or Darwin’s «warm little pond») water precipitated metal sulfide particles. Oceanic hydrothermal systems have a zonal structure reflected in ancient volcanogenic massive sulfide ore deposits. They reach many kilometers in diameter and date back to the Archean. Most abundant are pyrite (FeS2), chalcopyrite (CuFeS2), and sphalerite (ZnS), with additions of galena (PbS) and alabandite (MnS). ZnS and MnS have a unique ability to store radiation energy, e.g. from ultraviolet light. When replicating molecules were originating, the primordial atmospheric pressure was high enough (>100 bar) to precipitate near the Earth’s surface, and ultraviolet irradiation was 10 to 100 times more intense than now; hence the photosynthetic properties mediated by ZnS provided the right energy conditions for the synthesis of informational and metabolic molecules and the selection of photostable nucleobases.[224][226]
The Zn-world theory has been filled out with evidence for the ionic constitution of the interior of the first protocells. In 1926, the Canadian biochemist Archibald Macallum noted the resemblance of body fluids such as blood and lymph to seawater;[227] however, the inorganic composition of all cells differ from that of modern seawater, which led Mulkidjanian and colleagues to reconstruct the «hatcheries» of the first cells combining geochemical analysis with phylogenomic scrutiny of the inorganic ion requirements of modern cells. The authors conclude that ubiquitous, and by inference primordial, proteins and functional systems show affinity to and functional requirement for K+, Zn2+, Mn2+, and [PO
4]3−
. Geochemical reconstruction shows that this ionic composition could not have existed in the ocean but is compatible with inland geothermal systems. In the oxygen-depleted, CO2-dominated primordial atmosphere, the chemistry of water condensates near geothermal fields would resemble the internal milieu of modern cells. Therefore, precellular evolution may have taken place in shallow «Darwin ponds» lined with porous silicate minerals mixed with metal sulfides and enriched in K+, Zn2+, and phosphorus compounds.[228][229]
Homochirality[edit]

Many biomolecules, such as L-glutamic acid, are asymmetric, and occur in living systems in only one of the two possible forms, in the case of amino acids the left-handed form. Prebiotic chemistry would produce both forms, creating a puzzle for abiogenesis researchers.[230]
Homochirality is the geometric uniformity of materials composed of chiral (non-mirror-symmetric) units. Living organisms use molecules that have the same chirality (handedness): with almost no exceptions,[231] amino acids are left-handed while nucleotides and sugars are right-handed. Chiral molecules can be synthesized, but in the absence of a chiral source or a chiral catalyst, they are formed in a 50/50 (racemic) mixture of both forms. Known mechanisms for the production of non-racemic mixtures from racemic starting materials include: asymmetric physical laws, such as the electroweak interaction; asymmetric environments, such as those caused by circularly polarized light, quartz crystals, or the Earth’s rotation, statistical fluctuations during racemic synthesis,[230] and spontaneous symmetry breaking.[232][233][234] Once established, chirality would be selected for.[235] A small bias (enantiomeric excess) in the population can be amplified into a large one by asymmetric autocatalysis, such as in the Soai reaction.[236] In asymmetric autocatalysis, the catalyst is a chiral molecule, which means that a chiral molecule is catalyzing its own production. An initial enantiomeric excess, such as can be produced by polarized light, then allows the more abundant enantiomer to outcompete the other.[237] Homochirality may have started in outer space, as on the Murchison meteorite the amino acid L-alanine is more than twice as frequent as its D form, and L-glutamic acid is more than three times as abundant as its D counterpart.[238][239] Amino acids from meteorites show a left-handed bias, whereas sugars show a predominantly right-handed bias, as found in living organisms, suggesting an abiogenic origin of these compounds.[240]
See also[edit]
- Formamide-based prebiotic chemistry – Scientific efforts aimed at reconstructing the beginnings of life
- GADV-protein world hypothesis – A hypothetical stage of abiogenesis
- Genetic recombination – Production of offspring with combinations of traits that differ from those found in either parent
- Shadow biosphere – Hypothetical biosphere of Earth
References[edit]
Explanatory footnotes[edit]
- ^ The reactions are:
- FeS + H2S → FeS2 + 2H+ + 2e−
- FeS + H2S + CO2 → FeS2 + HCOOH
- ^ The reactions are:
Reaction 1: Fayalite + water → magnetite + aqueous silica + hydrogen- 3Fe2SiO4 + 2H2O → 2Fe3O4 + 3SiO2 + 2H2
Reaction 2: Forsterite + aqueous silica → serpentine
- 3Mg2SiO4 + SiO2 + 4H2O → 2Mg3Si2O5(OH)4
Reaction 3: Forsterite + water → serpentine + brucite
- 2Mg2SiO4 + 3H2O → Mg3Si2O5(OH)4 + Mg(OH)2
Reaction 3 describes the hydration of olivine with water only to yield serpentine and Mg(OH)2 (brucite). Serpentine is stable at high pH in the presence of brucite like calcium silicate hydrate, (C-S-H) phases formed along with portlandite (Ca(OH)2) in hardened Portland cement paste after the hydration of belite (Ca2SiO4), the artificial calcium equivalent of forsterite.
Analogy of reaction 3 with belite hydration in ordinary Portland cement: Belite + water → C-S-H phase + portlandite
- 2 Ca2SiO4 + 4 H2O → 3 CaO · 2 SiO2 · 3 H2O + Ca(OH)2
Citations[edit]
- ^ a b Walker, Sara I.; Packard, N.; Cody, G. D. (13 November 2017). «Re-conceptualizing the origins of life». Philosophical Transactions of the Royal Society A. 375 (2109): 20160337. Bibcode:2017RSPTA.37560337W. doi:10.1098/rsta.2016.0337. PMC 5686397. PMID 29133439.
- ^ a b c «NASA Astrobiology Strategy» (PDF). NASA. 2015. Archived from the original (PDF) on 22 December 2016. Retrieved 24 September 2017.
- ^ Trifonov, Edward N. (17 March 2011). «Vocabulary of Definitions of Life Suggests a Definition». Journal of Biomolecular Structure and Dynamics. 29 (2): 259–266. doi:10.1080/073911011010524992. PMID 21875147. S2CID 38476092.
- ^ Voytek, Mary A. (6 March 2021). «About Life Detection». NASA. Retrieved 8 March 2021.
- ^ a b Witzany, Guenther (2016). «Crucial steps to life: From chemical reactions to code using agents» (PDF). BioSystems. 140: 49–57. doi:10.1016/j.biosystems.2015.12.007. PMID 26723230. S2CID 30962295.
- ^ a b Howell, Elizabeth (8 December 2014). «How Did Life Become Complex, And Could It Happen Beyond Earth?». Astrobiology Magazine. Archived from the original on 15 February 2018. Retrieved 14 April 2022.
{{cite web}}: CS1 maint: unfit URL (link) - ^ Oparin, Aleksandr Ivanovich (2003) [1938]. The Origin of Life. Translated by Morgulis, Sergius (2 ed.). Mineola, New York: Courier. ISBN 978-0486495224.
- ^ a b Peretó, Juli (2005). «Controversies on the origin of life» (PDF). International Microbiology. 8 (1): 23–31. PMID 15906258. Archived from the original (PDF) on 24 August 2015. Retrieved 1 June 2015.
- ^ Compare: Scharf, Caleb; et al. (18 December 2015). «A Strategy for Origins of Life Research». Astrobiology. 15 (12): 1031–1042. Bibcode:2015AsBio..15.1031S. doi:10.1089/ast.2015.1113. PMC 4683543. PMID 26684503.
What do we mean by the origins of life (OoL)? … Since the early 20th century the phrase OoL has been used to refer to the events that occurred during the transition from non-living to living systems on Earth, i.e., the origin of terrestrial biology (Oparin, 1924; Haldane, 1929). The term has largely replaced earlier concepts such as abiogenesis (Kamminga, 1980; Fry, 2000).
- ^ a b c Weiss, M. C.; Sousa, F. L.; Mrnjavac, N.; Neukirchen, S.; Roettger, M.; Nelson-Sathi, S.; Martin, W.F. (2016). «The physiology and habitat of the last universal common ancestor» (PDF). Nature Microbiology. 1 (9): 16116. doi:10.1038/NMICROBIOL.2016.116. PMID 27562259. S2CID 2997255.
- ^ Tirard, Stephane (20 April 2015). Abiogenesis – Definition. Encyclopedia of Astrobiology. p. 1. doi:10.1007/978-3-642-27833-4_2-4. ISBN 978-3-642-27833-4.
Thomas Huxley (1825–1895) used the term abiogenesis in an important text published in 1870. He strictly made the difference between spontaneous generation, which he did not accept, and the possibility of the evolution of matter from inert to living, without any influence of life. … Since the end of the nineteenth century, evolutive abiogenesis means increasing complexity and evolution of matter from inert to living state in the abiotic context of evolution of primitive Earth.
- ^ Graham, Robert W. (February 1990). «Extraterrestrial Life in the Universe» (PDF). NASA (NASA Technical Memorandum 102363). Lewis Research Center, Cleveland, Ohio. Archived (PDF) from the original on 3 September 2014. Retrieved 2 June 2015.
- ^ Altermann 2009, p. xvii
- ^ Oparin 1953, p. vi
- ^ Warmflash, David; Warmflash, Benjamin (November 2005). «Did Life Come from Another World?». Scientific American. 293 (5): 64–71. Bibcode:2005SciAm.293e..64W. doi:10.1038/scientificamerican1105-64. PMID 16318028.
- ^ Yarus 2010, p. 47
- ^ Ward, Peter; Kirschvink, Joe (2015). A New History of Life: the radical discoveries about the origins and evolution of life on earth. Bloomsbury Press. pp. 39–40. ISBN 978-1608199105.
- ^ Sheldon 2005
- ^ Lennox 2001, pp. 229–258
- ^ a b Bernal 1967
- ^ Balme, D. M. (1962). «Development of Biology in Aristotle and Theophrastus: Theory of Spontaneous Generation». Phronesis. 7 (1–2): 91–104. doi:10.1163/156852862X00052.
- ^ Ross 1652
- ^ Dobell 1960
- ^ Bondeson 1999
- ^ Levine, R.; Evers, C. «The Slow Death of Spontaneous Generation (1668-1859)». Archived from the original on 26 April 2008. Retrieved 18 April 2013.
- ^ Oparin 1953, p. 196
- ^ Tyndall 1905, IV, XII (1876), XIII (1878)
- ^ Horneck, Gerda; Klaus, David M.; Mancinelli, Rocco L. (March 2010). «Space Microbiology». Microbiology and Molecular Biology Reviews. 74 (1): 121–156. Bibcode:2010MMBR…74..121H. doi:10.1128/MMBR.00016-09. PMC 2832349. PMID 20197502.
- ^ Wickramasinghe, Chandra (2011). «Bacterial morphologies supporting cometary panspermia: a reappraisal». International Journal of Astrobiology. 10 (1): 25–30. Bibcode:2011IJAsB..10…25W. CiteSeerX 10.1.1.368.4449. doi:10.1017/S1473550410000157. S2CID 7262449.
- ^ Rampelotto, P. H. (2010). «Panspermia: A promising field of research». In: Astrobiology Science Conference. Abs 5224.
- ^ Chang, Kenneth (12 September 2016). «Visions of Life on Mars in Earth’s Depths». The New York Times. Archived from the original on 12 September 2016. Retrieved 12 September 2016.
- ^ «Letter no. 7471, Charles Darwin to Joseph Dalton Hooker, 1 February (1871)». Darwin Correspondence Project. Retrieved 7 July 2020.
- ^ Priscu, John C. «Origin and Evolution of Life on a Frozen Earth». Arlington County, Virginia: National Science Foundation. Archived from the original on 18 December 2013. Retrieved 1 March 2014.
- ^ Marshall, Michael (11 November 2020). «Charles Darwin’s hunch about early life was probably right». BBC News. Retrieved 11 November 2020.
- ^ Bahadur, Krishna (1973). «Photochemical Formation of Self–sustaining Coacervates» (PDF). Proceedings of the Indian National Science Academy. 39 (4): 455–467. doi:10.1016/S0044-4057(75)80076-1. PMID 1242552. Archived from the original (PDF) on 19 October 2013.
- ^ Bahadur, Krishna (1975). «Photochemical Formation of Self-Sustaining Coacervates». Zentralblatt für Bakteriologie, Parasitenkunde, Infektionskrankheiten und Hygiene (Central Journal for Bacteriology, Parasitology, Infectious Diseases and Hygiene). 130 (3): 211–218. doi:10.1016/S0044-4057(75)80076-1. OCLC 641018092. PMID 1242552.
- ^ Bryson 2004, pp. 300–302
- ^ Bernal 1951
- ^ Martin, William F. (January 2003). «On the origins of cells: a hypothesis for the evolutionary transitions from abiotic geochemistry to chemoautotrophic prokaryotes, and from prokaryotes to nucleated cells». Phil. Trans. R. Soc. Lond. A. 358 (1429): 59–83. doi:10.1098/rstb.2002.1183. PMC 1693102. PMID 12594918.
- ^ Bernal, John Desmond (September 1949). «The Physical Basis of Life». Proceedings of the Physical Society, Section A. 62 (9): 537–558. Bibcode:1949PPSA…62..537B. doi:10.1088/0370-1298/62/9/301. S2CID 83754271.
- ^ Miller, Stanley L. (15 May 1953). «A Production of Amino Acids Under Possible Primitive Earth Conditions». Science. 117 (3046): 528–529. Bibcode:1953Sci…117..528M. doi:10.1126/science.117.3046.528. PMID 13056598.
- ^ Parker, Eric T.; Cleaves, Henderson J.; Dworkin, Jason P.; et al. (5 April 2011). «Primordial synthesis of amines and amino acids in a 1958 Miller H2S-rich spark discharge experiment». PNAS. 108 (14): 5526–5531. Bibcode:2011PNAS..108.5526P. doi:10.1073/pnas.1019191108. PMC 3078417. PMID 21422282.
- ^ Bernal 1967, p. 143
- ^ a b Cleaves, H. James; Chalmers, John H.; Lazcano, Antonio; et al. (April 2008). «A Reassessment of Prebiotic Organic Synthesis in Neutral Planetary Atmospheres». Origins of Life and Evolution of Biospheres. 38 (2): 105–115. Bibcode:2008OLEB…38..105C. doi:10.1007/s11084-007-9120-3. PMID 18204914. S2CID 7731172.
- ^ Chyba, Christopher F. (13 May 2005). «Rethinking Earth’s Early Atmosphere». Science. 308 (5724): 962–963. doi:10.1126/science.1113157. PMID 15890865. S2CID 93303848.
- ^ Barton et al. 2007, pp. 93–95
- ^ Bada & Lazcano 2009, pp. 56–57
- ^ Bada, Jeffrey L.; Lazcano, Antonio (2 May 2003). «Prebiotic Soup – Revisiting the Miller Experiment» (PDF). Science. 300 (5620): 745–746. doi:10.1126/science.1085145. PMID 12730584. S2CID 93020326. Archived (PDF) from the original on 4 March 2016. Retrieved 13 June 2015.
- ^ Marigo, Paola; et al. (6 July 2020). «Carbon star formation as seen through the non-monotonic initial–final mass relation». Nature Astronomy. 152 (11): 1102–1110. arXiv:2007.04163. Bibcode:2020NatAs…4.1102M. doi:10.1038/s41550-020-1132-1. S2CID 220403402.
- ^ «WMAP- Life in the Universe».
- ^ «Formation of Solar Systems: Solar Nebular Theory». University of Massachusetts Amherst. Retrieved 27 September 2019.
- ^ «Age of the Earth». United States Geological Survey. 9 July 2007. Archived from the original on 23 December 2005. Retrieved 10 January 2006.
- ^ Dalrymple 2001, pp. 205–221
- ^ Fesenkov 1959, p. 9
- ^ Kasting, James F. (12 February 1993). «Earth’s Early Atmosphere» (PDF). Science. 259 (5097): 920–926. Bibcode:1993Sci…259..920K. doi:10.1126/science.11536547. PMID 11536547. S2CID 21134564. Archived from the original (PDF) on 10 October 2015. Retrieved 28 July 2015.
- ^ Morse, John (September 1998). «Hadean Ocean Carbonate Geochemistry». Aquatic Geochemistry. 4 (3/4): 301–319. Bibcode:1998MinM…62.1027M. doi:10.1023/A:1009632230875. S2CID 129616933.
- ^ a b c d e f g h Follmann, Hartmut; Brownson, Carol (November 2009). «Darwin’s warm little pond revisited: from molecules to the origin of life». Naturwissenschaften. 96 (11): 1265–1292. Bibcode:2009NW…..96.1265F. doi:10.1007/s00114-009-0602-1. PMID 19760276. S2CID 23259886.
- ^ Morse, John W.; MacKenzie, Fred T. (1998). «Hadean Ocean Carbonate Geochemistry». Aquatic Geochemistry. 4 (3–4): 301–319. Bibcode:1998MinM…62.1027M. doi:10.1023/A:1009632230875. S2CID 129616933.
- ^ Wilde, Simon A.; Valley, John W.; Peck, William H.; Graham, Colin M. (11 January 2001). «Evidence from detrital zircons for the existence of continental crust and oceans on the Earth 4.4 Gyr ago» (PDF). Nature. 409 (6817): 175–178. Bibcode:2001Natur.409..175W. doi:10.1038/35051550. PMID 11196637. S2CID 4319774. Archived (PDF) from the original on 5 June 2015. Retrieved 3 June 2015.
- ^ Rosing, Minik T.; Bird, Dennis K.; Sleep, Norman H.; et al. (22 March 2006). «The rise of continents – An essay on the geologic consequences of photosynthesis». Palaeogeography, Palaeoclimatology, Palaeoecology. 232 (2–4): 99–113. Bibcode:2006PPP…232…99R. doi:10.1016/j.palaeo.2006.01.007. Archived (PDF) from the original on 14 July 2015. Retrieved 8 June 2015.
- ^ a b c Dodd, Matthew S.; Papineau, Dominic; Grenne, Tor; et al. (1 March 2017). «Evidence for early life in Earth’s oldest hydrothermal vent precipitates». Nature. 543 (7643): 60–64. Bibcode:2017Natur.543…60D. doi:10.1038/nature21377. PMID 28252057. Archived from the original on 8 September 2017. Retrieved 2 March 2017.
- ^ Gomes, Rodney; Levison, Hal F.; Tsiganis, Kleomenis; Morbidelli, Alessandro (26 May 2005). «Origin of the cataclysmic Late Heavy Bombardment period of the terrestrial planets». Nature. 435 (7041): 466–469. Bibcode:2005Natur.435..466G. doi:10.1038/nature03676. PMID 15917802.
- ^ Sleep, Norman H.; Zahnle, Kevin J.; Kasting, James F.; et al. (9 November 1989). «Annihilation of ecosystems by large asteroid impacts on early Earth». Nature. 342 (6246): 139–142. Bibcode:1989Natur.342..139S. doi:10.1038/342139a0. PMID 11536616. S2CID 1137852.
- ^ Chyba, Christopher; Sagan, Carl (9 January 1992). «Endogenous production, exogenous delivery and impact-shock synthesis of organic molecules: an inventory for the origins of life». Nature. 355 (6356): 125–132. Bibcode:1992Natur.355..125C. doi:10.1038/355125a0. PMID 11538392. S2CID 4346044.
- ^ Furukawa, Yoshihiro; Sekine, Toshimori; Oba, Masahiro; et al. (January 2009). «Biomolecule formation by oceanic impacts on early Earth». Nature Geoscience. 2 (1): 62–66. Bibcode:2009NatGe…2…62F. doi:10.1038/NGEO383.
- ^ Maher, Kevin A.; Stevenson, David J. (18 February 1988). «Impact frustration of the origin of life». Nature. 331 (6157): 612–614. Bibcode:1988Natur.331..612M. doi:10.1038/331612a0. PMID 11536595. S2CID 4284492.
- ^ Mann, Adam (24 January 2018). «Bashing holes in the tale of Earth’s troubled youth». Nature. 553 (7689): 393–395. Bibcode:2018Natur.553..393M. doi:10.1038/d41586-018-01074-6.
- ^ Davies 1999, p. 155
- ^ Bock & Goode 1996
- ^ Schopf, J. William; Kudryavtsev, Anatoliy B.; Czaja, Andrew D.; Tripathi, Abhishek B. (5 October 2007). «Evidence of Archean life: Stromatolites and microfossils». Precambrian Research. 158 (3–4): 141–155. Bibcode:2007PreR..158..141S. doi:10.1016/j.precamres.2007.04.009.
- ^ Schopf, J. William (29 June 2006). «Fossil evidence of Archaean life». Philosophical Transactions of the Royal Society B. 361 (1470): 869–885. doi:10.1098/rstb.2006.1834. PMC 1578735. PMID 16754604.
- ^ Raven & Johnson 2002, p. 68
- ^ Djokic, Tara; Van Kranendonk, Martin J.; Campbell, Kathleen A.; Walter, Malcolm R.; Ward, Colin R. (9 May 2017). «Earliest signs of life on land preserved in ca. 3.5 Gao hot spring deposits». Nature Communications. 8: 15263. Bibcode:2017NatCo…815263D. doi:10.1038/ncomms15263. PMC 5436104. PMID 28486437.
- ^ Schopf, J. William; Kitajima, Kouki; Spicuzza, Michael J.; Kudryavtsev, Anatolly B.; Valley, John W. (2017). «SIMS analyses of the oldest known assemblage of microfossils document their taxon-correlated carbon isotope compositions». PNAS. 115 (1): 53–58. Bibcode:2018PNAS..115…53S. doi:10.1073/pnas.1718063115. PMC 5776830. PMID 29255053.
- ^ Tyrell, Kelly April (18 December 2017). «Oldest fossils ever found show life on Earth began before 3.5 billion years ago». University of Wisconsin-Madison. Retrieved 18 December 2017.
- ^ Ohtomo, Yoko; Kakegawa, Takeshi; Ishida, Akizumi; et al. (January 2014). «Evidence for biogenic graphite in early Archaean Isua metasedimentary rocks». Nature Geoscience. 7 (1): 25–28. Bibcode:2014NatGe…7…25O. doi:10.1038/ngeo2025.
- ^ Noffke, Nora; Christian, Daniel; Wacey, David; Hazen, Robert M. (16 November 2013). «Microbially Induced Sedimentary Structures Recording an Ancient Ecosystem in the ca. 3.48 Gyo Dresser Formation, Pilbara, Western Australia». Astrobiology. 13 (12): 1103–1124. Bibcode:2013AsBio..13.1103N. doi:10.1089/ast.2013.1030. PMC 3870916. PMID 24205812.
- ^ Davies 1999
- ^ Hassenkam, T.; Andersson, M. P.; Dalby, K. N.; Mackenzie, D.M.A.; Rosing, M.T. (2017). «Elements of Eoarchean life trapped in mineral inclusions». Nature. 548 (7665): 78–81. Bibcode:2017Natur.548…78H. doi:10.1038/nature23261. PMID 28738409. S2CID 205257931.
- ^ O’Donoghue, James (21 August 2011). «Oldest reliable fossils show early life was a beach». New Scientist. 211: 13. doi:10.1016/S0262-4079(11)62064-2. Archived from the original on 30 June 2015.
- ^ Wacey, David; Kilburn, Matt R.; Saunders, Martin; et al. (October 2011). «Microfossils of sulphur-metabolizing cells in 3.4-billion-year-old rocks of Western Australia». Nature Geoscience. 4 (10): 698–702. Bibcode:2011NatGe…4..698W. doi:10.1038/ngeo1238.
- ^ Bell, Elizabeth A.; Boehnike, Patrick; Harrison, T. Mark; et al. (19 October 2015). «Potentially biogenic carbon preserved in a 4.1 billion-year-old zircon». PNAS. 112 (47): 14518–14521. Bibcode:2015PNAS..11214518B. doi:10.1073/pnas.1517557112. PMC 4664351. PMID 26483481. Early edition, published online before print.
- ^ Baumgartner, Rafael; Van Kranendonk, Martin; Wacey, David; et al. (2019). «Nano−porous pyrite and organic matter in 3.5-billion-year-old stromatolites record primordial life» (PDF). Geology. 47 (11): 1039–1043. Bibcode:2019Geo….47.1039B. doi:10.1130/G46365.1. S2CID 204258554.
- ^ Djokic, Tara; Van Kranendonk, Martin; Cambell, Kathleen; Walter, Malcolm (2017). «Earliest signs of life on land preserved in ca. 3.5 Ga hot spring deposits». Nature Communications. 3.
- ^ Landau, Elizabeth (12 October 2016). «Building Blocks of Life’s Building Blocks Come From Starlight». NASA. Archived from the original on 13 October 2016. Retrieved 13 October 2016.
- ^ a b Ehrenfreund, Pascale; Cami, Jan (December 2010). «Cosmic carbon chemistry: from the interstellar medium to the early Earth». Cold Spring Harbor Perspectives in Biology. 2 (12): a002097. doi:10.1101/cshperspect.a002097. PMC 2982172. PMID 20554702.
- ^ Geballe, Thomas R.; Najarro, Francisco; Figer, Donald F.; et al. (10 November 2011). «Infrared diffuse interstellar bands in the Galactic Centre region». Nature. 479 (7372): 200–202. arXiv:1111.0613. Bibcode:2011Natur.479..200G. doi:10.1038/nature10527. PMID 22048316. S2CID 17223339.
- ^ Klyce 2001
- ^ a b c d Hoover, Rachel (21 February 2014). «Need to Track Organic Nano-Particles Across the Universe? NASA’s Got an App for That». Ames Research Center. NASA. Archived from the original on 6 September 2015. Retrieved 22 June 2015.
- ^ Goncharuk, Vladislav V.; Zui, O. V. (February 2015). «Water and carbon dioxide as the main precursors of organic matter on Earth and in space». Journal of Water Chemistry and Technology. 37 (1): 2–3. doi:10.3103/S1063455X15010026. S2CID 97965067.
- ^ Abou Mrad, Ninette; Vinogradoff, Vassilissa; Duvernay, Fabrice; et al. (2015). «Laboratory experimental simulations: Chemical evolution of the organic matter from interstellar and cometary ice analogs». Bulletin de la Société Royale des Sciences de Liège. 84: 21–32. Bibcode:2015BSRSL..84…21A. Archived from the original on 13 April 2015. Retrieved 6 April 2015.
- ^ Oba, Yasuhiro; et al. (26 April 2022). «Identifying the wide diversity of extraterrestrial purine and pyrimidine nucleobases in carbonaceous meteorites». Nature Communications. 13 (2008): 2008. Bibcode:2022NatCo..13.2008O. doi:10.1038/s41467-022-29612-x. PMC 9042847. PMID 35473908. S2CID 248402205.
- ^ «‘Life chemical’ detected in comet». BBC News. London. 18 August 2009. Archived from the original on 25 May 2015. Retrieved 23 June 2015.
- ^ Thompson, William Reid; Murray, B. G.; Khare, Bishun Narain; Sagan, Carl (30 December 1987). «Coloration and darkening of methane clathrate and other ices by charged particle irradiation: Applications to the outer solar system». Journal of Geophysical Research. 92 (A13): 14933–14947. Bibcode:1987JGR….9214933T. doi:10.1029/JA092iA13p14933. PMID 11542127.
- ^ Goldman, Nir; Tamblyn, Isaac (20 June 2013). «Prebiotic Chemistry within a Simple Impacting Icy Mixture». Journal of Physical Chemistry A. 117 (24): 5124–5131. Bibcode:2013JPCA..117.5124G. doi:10.1021/jp402976n. PMID 23639050. S2CID 5144843.
- ^ «NASA Ames PAH IR Spectroscopic Database». NASA. Archived from the original on 29 June 2015. Retrieved 17 June 2015.
- ^ a b c Hudgins, Douglas M.; Bauschlicher, Charles W. Jr.; Allamandola, Louis J. (10 October 2005). «Variations in the Peak Position of the 6.2 μm Interstellar Emission Feature: A Tracer of N in the Interstellar Polycyclic Aromatic Hydrocarbon Population». The Astrophysical Journal. 632 (1): 316–332. Bibcode:2005ApJ…632..316H. CiteSeerX 10.1.1.218.8786. doi:10.1086/432495. S2CID 7808613.
- ^ a b c Des Marais, David J.; Allamandola, Louis J.; Sandford, Scott; et al. (2009). «Cosmic Distribution of Chemical Complexity». Ames Research Center. Mountain View, California: NASA. Archived from the original on 27 February 2014. Retrieved 24 June 2015.
- ^ a b Carey, Bjorn (18 October 2005). «Life’s Building Blocks ‘Abundant in Space’«. Space.com. Watsonville, California: Imaginova. Archived from the original on 26 June 2015. Retrieved 23 June 2015.
- ^ García-Hernández, Domingo. A.; Manchado, Arturo; García-Lario, Pedro; et al. (20 November 2010). «Formation of Fullerenes in H-Containing Planetary Nebulae». The Astrophysical Journal Letters. 724 (1): L39–L43. arXiv:1009.4357. Bibcode:2010ApJ…724L..39G. doi:10.1088/2041-8205/724/1/L39. S2CID 119121764.
- ^ d’Ischia, Marco; Manini, Paola; Moracci, Marco; et al. (21 August 2019). «Astrochemistry and Astrobiology: Materials Science in Wonderland?». International Journal of Molecular Sciences. 20 (17): 4079. doi:10.3390/ijms20174079. PMC 6747172. PMID 31438518.
- ^ Gudipati, Murthy S.; Yang, Rui (1 September 2012). «In-situ Probing of Radiation-induced Processing of Organics in Astrophysical Ice Analogs – Novel Laser Desorption Laser Ionization Time-of-flight Mass Spectroscopic Studies». The Astrophysical Journal Letters. 756 (1): L24. Bibcode:2012ApJ…756L..24G. doi:10.1088/2041-8205/756/1/L24. S2CID 5541727.
- ^ a b Gallori, Enzo (June 2011). «Astrochemistry and the origin of genetic material». Rendiconti Lincei. 22 (2): 113–118. doi:10.1007/s12210-011-0118-4. S2CID 96659714. «Paper presented at the Symposium ‘Astrochemistry: molecules in space and time’ (Rome, 4–5 November 2010), sponsored by Fondazione ‘Guido Donegani’, Accademia Nazionale dei Lincei.»
- ^ Martins, Zita (February 2011). «Organic Chemistry of Carbonaceous Meteorites». Elements. 7 (1): 35–40. doi:10.2113/gselements.7.1.35.
- ^ Martins, Zita; Botta, Oliver; Fogel, Marilyn L.; et al. (15 June 2008). «Extraterrestrial nucleobases in the Murchison meteorite». Earth and Planetary Science Letters. 270 (1–2): 130–136. arXiv:0806.2286. Bibcode:2008E&PSL.270..130M. doi:10.1016/j.epsl.2008.03.026. S2CID 14309508.
- ^ Callahan, Michael P.; Smith, Karen E.; Cleaves, H. James, II; et al. (23 August 2011). «Carbonaceous meteorites contain a wide range of extraterrestrial nucleobases». PNAS. 108 (34): 13995–13998. Bibcode:2011PNAS..10813995C. doi:10.1073/pnas.1106493108. PMC 3161613. PMID 21836052.
- ^ Steigerwald, John (8 August 2011). «NASA Researchers: DNA Building Blocks Can Be Made in Space». Goddard Space Flight Center. NASA. Archived from the original on 23 June 2015. Retrieved 23 June 2015.
- ^ Kwok, Sun; Zhang, Yong (3 November 2011). «Mixed aromatic–aliphatic organic nanoparticles as carriers of unidentified infrared emission features». Nature. 479 (7371): 80–83. Bibcode:2011Natur.479…80K. doi:10.1038/nature10542. PMID 22031328. S2CID 4419859.
- ^ Jørgensen, Jes K.; Favre, Cécile; Bisschop, Suzanne E.; et al. (2012). «Detection of the simplest sugar, glycolaldehyde, in a solar-type protostar with ALMA» (PDF). The Astrophysical Journal Letters. 757 (1): L4. arXiv:1208.5498. Bibcode:2012ApJ…757L…4J. doi:10.1088/2041-8205/757/1/L4. S2CID 14205612. Archived (PDF) from the original on 24 September 2015. Retrieved 23 June 2015.
- ^ Furukawa, Yoshihiro; Chikaraishi, Yoshito; Ohkouchi, Naohiko; et al. (13 November 2019). «Extraterrestrial ribose and other sugars in primitive meteorites». PNAS. 116 (49): 24440–24445. Bibcode:2019PNAS..11624440F. doi:10.1073/pnas.1907169116. PMC 6900709. PMID 31740594.
- ^ Oró, Joan; Kimball, Aubrey P. (February 1962). «Synthesis of purines under possible primitive earth conditions: II. Purine intermediates from hydrogen cyanide». Archives of Biochemistry and Biophysics. 96 (2): 293–313. doi:10.1016/0003-9861(62)90412-5. PMID 14482339.
- ^ Cleaves II, Henderson (2010). «The origin of the biologically coded amino acids». Journal of Theoretical Biology. 263 (4): 490–498. Bibcode:2010JThBi.263..490C. doi:10.1016/j.jtbi.2009.12.014. PMID 20034500.
- ^ Breslow, R. (1959). «On the Mechanism of the Formose Reaction». Tetrahedron Letters. 1 (21): 22–26. doi:10.1016/S0040-4039(01)99487-0.
- ^ Oró, Joan (16 September 1961). «Mechanism of Synthesis of Adenine from Hydrogen Cyanide under Possible Primitive Earth Conditions». Nature. 191 (4794): 1193–1194. Bibcode:1961Natur.191.1193O. doi:10.1038/1911193a0. PMID 13731264. S2CID 4276712.
- ^ a b Saladino, Raffaele; Crestini, Claudia; Pino, Samanta; et al. (March 2012). «Formamide and the origin of life» (PDF). Physics of Life Reviews. 9 (1): 84–104. Bibcode:2012PhLRv…9…84S. doi:10.1016/j.plrev.2011.12.002. hdl:2108/85168. PMID 22196896.
- ^ a b Saladino, Raffaele; Botta, Giorgia; Pino, Samanta; et al. (July 2012). «From the one-carbon amide formamide to RNA all the steps are prebiotically possible». Biochimie. 94 (7): 1451–1456. doi:10.1016/j.biochi.2012.02.018. hdl:11573/515604. PMID 22738728.
- ^ Marlaire, Ruth, ed. (3 March 2015). «NASA Ames Reproduces the Building Blocks of Life in Laboratory». Ames Research Center. NASA. Archived from the original on 5 March 2015. Retrieved 5 March 2015.
- ^ Ferus, Martin; Nesvorný, David; Šponer, Jiří; et al. (2015). «High-energy chemistry of formamide: A unified mechanism of nucleobase formation». PNAS. 112 (3): 657–662. Bibcode:2015PNAS..112..657F. doi:10.1073/pnas.1412072111. PMC 4311869. PMID 25489115.
- ^ Basile, Brenda; Lazcano, Antonio; Oró, Joan (1984). «Prebiotic syntheses of purines and pyrimidines». Advances in Space Research. 4 (12): 125–131. Bibcode:1984AdSpR…4l.125B. doi:10.1016/0273-1177(84)90554-4. PMID 11537766.
- ^ Orgel, Leslie E. (August 2004). «Prebiotic Adenine Revisited: Eutectics and Photochemistry». Origins of Life and Evolution of Biospheres. 34 (4): 361–369. Bibcode:2004OLEB…34..361O. doi:10.1023/B:ORIG.0000029882.52156.c2. PMID 15279171. S2CID 4998122.
- ^ Robertson, Michael P.; Miller, Stanley L. (29 June 1995). «An efficient prebiotic synthesis of cytosine and uracil». Nature. 375 (6534): 772–774. Bibcode:1995Natur.375..772R. doi:10.1038/375772a0. PMID 7596408. S2CID 4351012.
- ^ Fox, Douglas (February 2008). «Did Life Evolve in Ice?». Discover. Archived from the original on 30 June 2008. Retrieved 3 July 2008.
- ^ Levy, Matthew; Miller, Stanley L.; Brinton, Karen; Bada, Jeffrey L. (June 2000). «Prebiotic Synthesis of Adenine and Amino Acids Under Europa-like Conditions». Icarus. 145 (2): 609–613. Bibcode:2000Icar..145..609L. doi:10.1006/icar.2000.6365. PMID 11543508.
- ^ Menor-Salván, César; Ruiz-Bermejo, Marta; Guzmán, Marcelo I.; et al. (20 April 2009). «Synthesis of Pyrimidines and Triazines in Ice: Implications for the Prebiotic Chemistry of Nucleobases». Chemistry: A European Journal. 15 (17): 4411–4418. doi:10.1002/chem.200802656. PMID 19288488.
- ^ Roy, Debjani; Najafian, Katayoun; von Ragué Schleyer, Paul (30 October 2007). «Chemical evolution: The mechanism of the formation of adenine under prebiotic conditions». PNAS. 104 (44): 17272–17277. Bibcode:2007PNAS..10417272R. doi:10.1073/pnas.0708434104. PMC 2077245. PMID 17951429.
- ^ Lancet, Doron (30 December 2014). «Systems Prebiology-Studies of the origin of Life». The Lancet Lab. Rehovot, Israel: Department of Molecular Genetics; Weizmann Institute of Science. Archived from the original on 26 June 2015. Retrieved 26 June 2015.
- ^ Segré, Daniel; Ben-Eli, Dafna; Deamer, David W.; Lancet, Doron (February 2001). «The Lipid World» (PDF). Origins of Life and Evolution of Biospheres. 31 (1–2): 119–145. Bibcode:2001OLEB…31..119S. doi:10.1023/A:1006746807104. PMID 11296516. S2CID 10959497. Archived (PDF) from the original on 26 June 2015.
- ^ a b c Chen, Irene A.; Walde, Peter (July 2010). «From Self-Assembled Vesicles to Protocells». Cold Spring Harbor Perspectives in Biology. 2 (7): a002170. doi:10.1101/cshperspect.a002170. PMC 2890201. PMID 20519344.
- ^ Eigen, Manfred; Schuster, Peter (November 1977). «The Hypercycle. A Principle of Natural Self-Organization. Part A: Emergence of the Hypercycle» (PDF). Naturwissenschaften. 64 (11): 541–65. Bibcode:1977NW…..64..541E. doi:10.1007/bf00450633. PMID 593400. S2CID 42131267. Archived from the original (PDF) on 3 March 2016.
- Eigen, Manfred; Schuster, Peter (1978). «The Hypercycle. A Principle of Natural Self-Organization. Part B: The Abstract Hypercycle» (PDF). Naturwissenschaften. 65 (1): 7–41. Bibcode:1978NW…..65….7E. doi:10.1007/bf00420631. S2CID 1812273. Archived from the original (PDF) on 3 March 2016.
- Eigen, Manfred; Schuster, Peter (July 1978). «The Hypercycle. A Principle of Natural Self-Organization. Part C: The Realistic Hypercycle» (PDF). Naturwissenschaften. 65 (7): 341–369. Bibcode:1978NW…..65..341E. doi:10.1007/bf00439699. S2CID 13825356. Archived from the original (PDF) on 16 June 2016.
- ^ Markovitch, Omer; Lancet, Doron (Summer 2012). «Excess Mutual Catalysis Is Required for Effective Evolvability». Artificial Life. 18 (3): 243–266. doi:10.1162/artl_a_00064. PMID 22662913. S2CID 5236043.
- ^ Tessera, Marc (2011). «Origin of Evolution versus Origin of Life: A Shift of Paradigm». International Journal of Molecular Sciences. 12 (6): 3445–3458. doi:10.3390/ijms12063445. PMC 3131571. PMID 21747687. Special Issue: «Origin of Life 2011»
- ^ Onsager, Lars (1931). «Reciprocal Relations in Irreversible Processes I and II». Physical Review. 37 (4): 405. Bibcode:1931PhRv…37..405O. doi:10.1103/PhysRev.37.405.
- ^ Onsager, Lars (1931). «Reciprocal Relations in Irreversible Processes I and II». Physical Review (38): 2265. doi:10.1103/PhysRev.38.2265.
- ^ Prigogine, Ilya (1967). An Introduction to the Thermodynamics of Irreversible Processes. New York: Wiley.
- ^ «Exploring Life’s Origins: Protocells». Exploring Life’s Origins: A Virtual Exhibit. Arlington County, Virginia: National Science Foundation. Archived from the original on 28 February 2014. Retrieved 18 March 2014.
- ^ a b c Chen, Irene A. (8 December 2006). «The Emergence of Cells During the Origin of Life». Science. 314 (5805): 1558–1559. doi:10.1126/science.1137541. PMID 17158315.
- ^ Zimmer, Carl (26 June 2004). «What Came Before DNA?». Discover. Archived from the original on 19 March 2014.
- ^ Shapiro, Robert (June 2007). «A Simpler Origin for Life». Scientific American. 296 (6): 46–53. Bibcode:2007SciAm.296f..46S. doi:10.1038/scientificamerican0607-46. PMID 17663224. Archived from the original on 14 June 2015.
- ^ Chang 2007
- ^ a b c d Lane, Nick (2015). The Vital Question: Why Is Life The Way It Is?. Profile Books. pp. 129–140. ISBN 978-1781250365.
- ^ Sharov, Alexei A.; Gordon, Richard (2018). «Life Before Earth». Habitability of the Universe Before Earth: Life Before Earth. Astrobiology Exploring Life on Earth and Beyond. Academic Press. pp. 265–296. doi:10.1016/B978-0-12-811940-2.00011-3. ISBN 9780128119402. S2CID 117048600.
- ^ Ladyman, J.; Lambert, J.; Weisner, K. B. (2013). «What is a Complex System?». European Journal of the Philosophy of Science. 3: 33–67. doi:10.1007/s13194-012-0056-8. S2CID 18787276.
- ^ Esposito, M.; Lindenberg, Katja; Van den Broeck, C. (2010). «Entropy production as correlation between system and reservoir». New Journal of Physics. 12 (1): 013013. arXiv:0908.1125. Bibcode:2010NJPh…12a3013E. doi:10.1088/1367-2630/12/1/013013. S2CID 26657293.
- ^ Bar-Nun, A.; Bar-Nun, N.; Bauer, S. H.; Sagan, Carl (24 April 1970). «Shock Synthesis of Amino Acids in Simulated Primitive Environments». Science. 168 (3930): 470–473. Bibcode:1970Sci…168..470B. doi:10.1126/science.168.3930.470. PMID 5436082. S2CID 42467812.
- ^ Anbar, Michael (27 September 1968). «Cavitation during Impact of Liquid Water on Water: Geochemical Implications». Science. 161 (3848): 1343–1344. Bibcode:1968Sci…161.1343A. doi:10.1126/science.161.3848.1343. PMID 17831346.
- ^ Dharmarathne, Leena; Grieser, Franz (7 January 2016). «Formation of Amino Acids on the Sonolysis of Aqueous Solutions Containing Acetic Acid, Methane, or Carbon Dioxide, in the Presence of Nitrogen Gas». The Journal of Physical Chemistry A. 120 (2): 191–199. Bibcode:2016JPCA..120..191D. doi:10.1021/acs.jpca.5b11858. PMID 26695890.
- ^ Patehebieke, Yeersen; Zhao, Ze-Run; Wang, Su; Xu, Hao-Xing; Chen, Qian-Qian; Wang, Xiao (2021). «Cavitation as a plausible driving force for the prebiotic formation of N9 purine nucleosides». Cell Reports Physical Science. 2 (3): 100375. Bibcode:2021CRPS….200375P. doi:10.1016/j.xcrp.2021.100375. S2CID 233662126.
- ^ Kalson, Natan-Haim; Furman, David; Zeiri, Yehuda (11 September 2017). «Cavitation-Induced Synthesis of Biogenic Molecules on Primordial Earth». ACS Central Science. 3 (9): 1041–1049. doi:10.1021/acscentsci.7b00325. PMC 5620973. PMID 28979946. S2CID 21409351.
- ^ Muller, Anthonie W. J. (1995). «Were the first organisms heat engines? A new model for biogenesis and the early evolution of biological energy conversion». Progress in Biophysics and Molecular Biology. 63 (2): 193–231. doi:10.1016/0079-6107(95)00004-7. PMID 7542789.
- ^ Muller, Anthonie W. J.; Schulze-Makuch, Dirk (2006). «Thermal energy and the origin of life». Origins of Life and Evolution of Biospheres. 36 (2): 77–189. Bibcode:2006OLEB…36..177M. doi:10.1007/s11084-005-9003-4. PMID 16642267. S2CID 22179552.
- ^ Junge, Wolfgang; Nelson, Nathan (2 June 2015). «ATP Synthase». Annual Review of Biochemistry. 84 (1): 631–657. doi:10.1146/annurev-biochem-060614-034124. PMID 25839341.
- ^ a b c Damer, Bruce; Deamer, David (1 April 2020). «The Hot Spring Hypothesis for an Origin of Life». Astrobiology. 20 (4): 429–452. Bibcode:2020AsBio..20..429D. doi:10.1089/ast.2019.2045. ISSN 1531-1074. PMC 7133448. PMID 31841362.
- ^ Benner, S. A.; Bell, E. A.; Biondi, E.; Brasser, R.; Carell, T.; Kim, H.-J.; Mojzsis, S. J.; Omran, A.; Pasek, M. A.; Trail, D. (2020). «When Did Life Likely Emerge on Earth in an RNA-First Process?». ChemSystemsChem. 2 (2). doi:10.1002/syst.201900035.
- ^ * Copley, Shelley D.; Smith, Eric; Morowitz, Harold J. (December 2007). «The origin of the RNA world: Co-evolution of genes and metabolism» (PDF). Bioorganic Chemistry. 35 (6): 430–443. doi:10.1016/j.bioorg.2007.08.001. PMID 17897696. Archived (PDF) from the original on 5 September 2013. Retrieved 8 June 2015.
The proposal that life on Earth arose from an RNA world is widely accepted.
- Orgel, Leslie E. (April 2003). «Some consequences of the RNA world hypothesis». Origins of Life and Evolution of Biospheres. 33 (2): 211–218. Bibcode:2003OLEB…33..211O. doi:10.1023/A:1024616317965. PMID 12967268. S2CID 32779859.
It now seems very likely that our familiar DNA/RNA/protein world was preceded by an RNA world…
- Robertson & Joyce 2012: «There is now strong evidence indicating that an RNA World did indeed exist before DNA- and protein-based life.»
- Neveu, Kim & Benner 2013: «[The RNA world’s existence] has broad support within the community today.»
- Orgel, Leslie E. (April 2003). «Some consequences of the RNA world hypothesis». Origins of Life and Evolution of Biospheres. 33 (2): 211–218. Bibcode:2003OLEB…33..211O. doi:10.1023/A:1024616317965. PMID 12967268. S2CID 32779859.
- ^ a b c Robertson, Michael P.; Joyce, Gerald F. (May 2012). «The origins of the RNA world». Cold Spring Harbor Perspectives in Biology. 4 (5): a003608. doi:10.1101/cshperspect.a003608. PMC 3331698. PMID 20739415.
- ^ a b c Cech, Thomas R. (July 2012). «The RNA Worlds in Context». Cold Spring Harbor Perspectives in Biology. 4 (7): a006742. doi:10.1101/cshperspect.a006742. PMC 3385955. PMID 21441585.
- ^ Pearce, Ben K. D.; Pudritz, Ralph E.; Semenov, Dmitry A.; Henning, Thomas K. (24 October 2017). «Origin of the RNA world: The fate of nucleobases in warm little ponds». PNAS. 114 (43): 11327–11332. arXiv:1710.00434. Bibcode:2017PNAS..11411327P. doi:10.1073/pnas.1710339114. PMC 5664528. PMID 28973920.
- ^ Yarus, Michael (April 2011). «Getting Past the RNA World: The Initial Darwinian Ancestor». Cold Spring Harbor Perspectives in Biology. 3 (4): a003590. doi:10.1101/cshperspect.a003590. PMC 3062219. PMID 20719875.
- ^ Voet & Voet 2004, p. 29
- ^ Fox, George.E. (9 June 2010). «Origin and evolution of the ribosome». Cold Spring Harbor Perspectives in Biology. 2 (9(a003483)): a003483. doi:10.1101/cshperspect.a003483. PMC 2926754. PMID 20534711.
- ^ Neveu, Marc; Kim, Hyo-Joong; Benner, Steven A. (22 April 2013). «The ‘Strong’ RNA World Hypothesis: Fifty Years Old». Astrobiology. 13 (4): 391–403. Bibcode:2013AsBio..13..391N. doi:10.1089/ast.2012.0868. PMID 23551238.
- ^ Gilbert, Walter (20 February 1986). «Origin of life: The RNA world». Nature. 319 (6055): 618. Bibcode:1986Natur.319..618G. doi:10.1038/319618a0. S2CID 8026658.
- ^ Orgel, Leslie E. (October 1994). «The origin of life on Earth». Scientific American. 271 (4): 76–83. Bibcode:1994SciAm.271d..76O. doi:10.1038/scientificamerican1094-76. PMID 7524147.
- ^ Lincoln, Tracey A.; Joyce, Gerald F. (27 February 2009). «Self-Sustained Replication of an RNA Enzyme». Science. 323 (5918): 1229–1232. Bibcode:2009Sci…323.1229L. doi:10.1126/science.1167856. PMC 2652413. PMID 19131595.
- ^ Joyce, Gerald F. (2009). «Evolution in an RNA world». Cold Spring Harbor Perspectives in Biology. 74 (Evolution: The Molecular Landscape): 17–23. doi:10.1101/sqb.2009.74.004. PMC 2891321. PMID 19667013.
- ^ Szostak, Jack W. (5 February 2015). «The Origins of Function in Biological Nucleic Acids, Proteins, and Membranes». Chevy Chase, Maryland: Howard Hughes Medical Institute. Archived from the original on 14 July 2015. Retrieved 16 June 2015.
- ^ Bernstein, Harris; Byerly, Henry C.; Hopf, Frederick A.; et al. (June 1983). «The Darwinian Dynamic». The Quarterly Review of Biology. 58 (2): 185–207. doi:10.1086/413216. JSTOR 2828805. S2CID 83956410.
- ^ Michod 1999
- ^ Palasek, Stan (23 May 2013). «Primordial RNA Replication and Applications in PCR Technology». arXiv:1305.5581v1 [q-bio.BM].
- ^ Vlassov, Alexander V.; Kazakov, Sergei A.; Johnston, Brian H.; et al. (August 2005). «The RNA World on Ice: A New Scenario for the Emergence of RNA Information». Journal of Molecular Evolution. 61 (2): 264–273. Bibcode:2005JMolE..61..264V. doi:10.1007/s00239-004-0362-7. PMID 16044244. S2CID 21096886.
- ^ Nussinov, Mark D.; Otroshchenko, Vladimir A.; Santoli, Salvatore (1997). «The emergence of the non-cellular phase of life on the fine-grained clayish particles of the early Earth’s regolith». BioSystems. 42 (2–3): 111–118. doi:10.1016/S0303-2647(96)01699-1. PMID 9184757.
- ^ Kühnlein, Alexandra; Lanzmich, Simon A.; Brun, Dieter (2 March 2021). «tRNA sequences can assemble into a replicator». eLife. 10. doi:10.7554/eLife.63431. PMC 7924937. PMID 33648631.
- ^ Noller, Harry F. (April 2012). «Evolution of protein synthesis from an RNA world». Cold Spring Harbor Perspectives in Biology. 4 (4): a003681. doi:10.1101/cshperspect.a003681. PMC 3312679. PMID 20610545.
- ^ Koonin, Eugene V. (31 May 2007). «The cosmological model of eternal inflation and the transition from chance to biological evolution in the history of life». Biology Direct. 2: 15. doi:10.1186/1745-6150-2-15. PMC 1892545. PMID 17540027.
- ^ Boone, David R.; Castenholz, Richard W.; Garrity, George M., eds. (2001). The Archaea and the Deeply Branching and Phototrophic Bacteria. Bergey’s Manual of Systematic Bacteriology. Springer. ISBN 978-0-387-21609-6. Archived from the original on 25 December 2014.[page needed]
- ^ Woese, C. R.; Fox, G. E. (1977). «Phylogenetic structure of the prokaryotic domain: the primary kingdoms». PNAS. 7 (11): 5088–5090. Bibcode:1977PNAS…74.5088W. doi:10.1073/pnas.74.11.5088. PMC 432104. PMID 270744.
- ^ Valas, R. E.; Bourne, P. E. (2011). «The origin of a derived superkingdom: how a gram-positive bacterium crossed the desert to become an archaeon». Biology Direct. 6: 16. doi:10.1186/1745-6150-6-16. PMC 3056875. PMID 21356104.
- ^ Cavalier-Smith, Thomas (2006). «Rooting the tree of life by transition analyses». Biology Direct. 1: 19. doi:10.1186/1745-6150-1-19. PMC 1586193. PMID 16834776.
- ^ «Early life liked it hot». Nature. 535 (7613): 468. 2016. doi:10.1038/535468b. S2CID 49905802.
- ^ Gogarten, Johann Peter; Deamer, David (25 November 2016). «Is LUCA a thermophilic progenote?». Nature Microbiology. 1 (12): 16229. doi:10.1038/nmicrobiol.2016.229. ISSN 2058-5276. PMID 27886195. S2CID 205428194.
- ^ Catchpole, Ryan; Forterre, Patrick (2019). «The evolution of Reverse Gyrase suggests a non-hyperthermophilic Last Universal Common Ancestor». Molecular Biology and Evolution. 36 (12): 2737–2747. doi:10.1093/molbev/msz180. PMC 6878951. PMID 31504731.
- ^ Berkemer, Sarah J.; McGlynn, Shawn E (8 August 2020). «A New Analysis of Archaea–Bacteria Domain Separation: Variable Phylogenetic Distance and the Tempo of Early Evolution». Molecular Biology and Evolution. 37 (8): 2332–2340. doi:10.1093/molbev/msaa089. PMC 7403611. PMID 32316034.
- ^ Hoffmann, Geoffrey W. (25 June 1974). «On the origin of the genetic code and the stability of the translation apparatus». Journal of Molecular Biology. 86 (2): 349–362. doi:10.1016/0022-2836(74)90024-2. PMID 4414916.
- ^ Orgel, Leslie E. (April 1963). «The Maintenance of the Accuracy of Protein Synthesis and its Relevance to Ageing». PNAS. 49 (4): 517–521. Bibcode:1963PNAS…49..517O. doi:10.1073/pnas.49.4.517. PMC 299893. PMID 13940312.
- ^ Hoffmann, Geoffrey W. (October 1975). «The Stochastic Theory of the Origin of the Genetic Code». Annual Review of Physical Chemistry. 26: 123–144. Bibcode:1975ARPC…26..123H. doi:10.1146/annurev.pc.26.100175.001011.
- ^ Brasier, M. D. (2012). Secret Chambers: The Inside Story of Cells and Complex Life. Oxford University Press. p. 298.
- ^ Ward, Peter & Kirschvink, Joe, op cit, p. 42
- ^ a b Colín-García, M.; Heredia, A.; Cordero, G.; et al. (2016). «Hydrothermal vents and prebiotic chemistry: a review». Boletín de la Sociedad Geológica Mexicana. 68 (3): 599–620. doi:10.18268/BSGM2016v68n3a13. Archived from the original on 18 August 2017.
- ^ Schirber, Michael (24 June 2014). «Hydrothermal Vents Could Explain Chemical Precursors to Life». NASA Astrobiology: Life in the Universe. NASA. Archived from the original on 29 November 2014. Retrieved 19 June 2015.
- ^ a b Martin, William; Russell, Michael J. (29 January 2003). «On the origins of cells: a hypothesis for the evolutionary transitions from abiotic geochemistry to chemoautotrophic prokaryotes, and from prokaryotes to nucleated cells». Philosophical Transactions of the Royal Society B. 358 (1429): 59–83, discussion 83–85. doi:10.1098/rstb.2002.1183. PMC 1693102. PMID 12594918.
- ^ a b Lane 2009
- ^ Usher, Oli (27 April 2015). «Chemistry of seabed’s hot vents could explain emergence of life» (Press release). University College London. Archived from the original on 20 June 2015. Retrieved 19 June 2015.
- ^ Roldan, Alberto; Hollingsworth, Nathan; Roffey, Anna; et al. (May 2015). «Bio-inspired CO2 conversion by iron sulfide catalysts under sustainable conditions». Chemical Communications. 51 (35): 7501–7504. doi:10.1039/C5CC02078F. PMID 25835242. Archived from the original on 20 June 2015. Retrieved 19 June 2015.
- ^ Baross, J. A.; Hoffman, S. E. (1985). «Submarine hydrothermal vents and associated gradient environments as sites for the origin and evolution of life». Origins of Life and Evolution of Biospheres. 15 (4): 327–345. Bibcode:1985OrLi…15..327B. doi:10.1007/bf01808177. S2CID 4613918.
- ^ Russell, M. J.; Hall, A. J. (1997). «The emergence of life from iron monosulphide bubbles at a submarine hydrothermal redox and pH front». Journal of the Geological Society. 154 (3): 377–402. Bibcode:1997JGSoc.154..377R. doi:10.1144/gsjgs.154.3.0377. PMID 11541234. S2CID 24792282.
- ^ Amend, J. P.; LaRowe, D. E.; McCollom, T. M.; Shock, E. L. (2013). «The energetics of organic synthesis inside and outside the cell». Philosophical Transactions of the Royal Society B. 368 (1622): 20120255. doi:10.1098/rstb.2012.0255. PMC 3685458. PMID 23754809.
- ^ Shock, E. L.; Boyd, E. S. (2015). «Geomicrobiology and microbial geochemistry:principles of geobiochemistry». Elements. 11: 389–394. doi:10.2113/gselements.11.6.395.
- ^ Martin, W.; Russell, M. J. (2007). «On the origin of biochemistry at an alkaline hydrothermal vent». Philosophical Transactions of the Royal Society B. 362 (1486): 1887–1925. doi:10.1098/rstb.2006.1881. PMC 2442388. PMID 17255002.
- ^ Weiss, Madeline C.; Sousa, Filipa L.; Mrnjavac, Natalia; Neukirchen, Sinje; Roettger, Mayo; Nelson-Sathi, Shijulal; Martin, William F. (25 July 2016). «The physiology and habitat of the last universal common ancestor» (PDF). Nature Microbiology. 1 (9): 16116. doi:10.1038/nmicrobiol.2016.116. PMID 27562259. S2CID 2997255.
- ^ Lane, Nick; Martin, William F. (21 December 2012). «The Origin of Membrane Bioenergetics». Cell. 151 (7): 1406–1416. doi:10.1016/j.cell.2012.11.050. ISSN 0092-8674. PMID 23260134. S2CID 15028935.
- ^ Baaske, Philipp; Weinert, Franz M.; Duhr, Stefan; Lemke, Kono H.; Russell, Michael J.; Braun, Dieter (29 May 2007). «Extreme accumulation of nucleotides in simulated hydrothermal pore systems». Proceedings of the National Academy of Sciences. 104 (22): 9346–9351. doi:10.1073/pnas.0609592104. ISSN 0027-8424. PMC 1890497. PMID 17494767.
- ^ a b c Mulkidjanian, Armen Y.; Bychkov, Andrew Yu.; Dibrova, Daria V.; Galperin, Michael Y.; Koonin, Eugene V. (3 April 2012). «Origin of first cells at terrestrial, anoxic geothermal fields». Proceedings of the National Academy of Sciences. 109 (14): E821-30. Bibcode:2012PNAS..109E.821M. doi:10.1073/pnas.1117774109. PMC 3325685. PMID 22331915.
- ^ Chandru, Kuhan; Guttenberg, Nicholas; Giri, Chaitanya; et al. (31 May 2018). «Simple prebiotic synthesis of high diversity dynamic combinatorial polyester libraries». Communications Chemistry. 1 (1). doi:10.1038/s42004-018-0031-1.
- ^ Forsythe, Jay G.; Yu, Sheng-Sheng; Mamajanov, Irena; et al. (17 August 2015). «Ester-Mediated Amide Bond Formation Driven by Wet–Dry Cycles: A Possible Path to Polypeptides on the Prebiotic Earth». Angewandte Chemie International Edition in English. 54 (34): 9871–9875. doi:10.1002/anie.201503792. PMC 4678426. PMID 26201989.
- ^ Patel, Bhavesh H.; Percivalle, Claudia; Ritson, Dougal J.; Duffy, Colm. D.; Sutherland, John D. (16 March 2015). «Common origins of RNA, protein and lipid precursors in a cyanosulfidic protometabolism». Nature Chemistry. 7 (4): 301–307. Bibcode:2015NatCh…7..301P. doi:10.1038/nchem.2202. ISSN 1755-4330. PMC 4568310. PMID 25803468.
- ^ Deamer, David (10 February 2021). «Where Did Life Begin? Testing Ideas in Prebiotic Analogue Conditions». Life. 11 (2): 134. doi:10.3390/life11020134. PMC 7916457. PMID 33578711.
- ^ Cairns-Smith, Graham (2 September 1982). Genetic Takeover and the Mineral Origins of Life. Cambridge: Cambridge University Press. ISBN 0-521-23312-7. OCLC 7875600.
- ^ Dawkins 1996, pp. 148–161
- ^ Huang, Wenhua; Ferris, James P. (12 July 2006). «One-Step, Regioselective Synthesis of up to 50-mers of RNA Oligomers by Montmorillonite Catalysis». Journal of the American Chemical Society. 128 (27): 8914–8919. doi:10.1021/ja061782k. PMID 16819887.
- ^ Subramaniam, Anand Bala; Wan, Jiandi; Gopinath, Arvind; Stone, Howard A. (2011). «Semi-permeable vesicles composed of natural clay» (PDF). Soft Matter. 7 (6): 2600–2612. arXiv:1011.4711. Bibcode:2011SMat….7.2600S. doi:10.1039/c0sm01354d. S2CID 52253528.
- ^ Hartman, Hyman (1998). «Photosynthesis and the Origin of Life». Origins of Life and Evolution of Biospheres. 28 (4–6): 515–521. Bibcode:1998OLEB…28..515H. doi:10.1023/A:1006548904157. PMID 11536891. S2CID 2464.
- ^ Yue-Ching Ho, Eugene (July–September 1990). «Evolutionary Epistemology and Sir Karl Popper’s Latest Intellectual Interest: A First-Hand Report». Intellectus. 15: 1–3. OCLC 26878740. Archived from the original on 11 March 2012.
- ^ Popper, Karl R. (29 March 1990). «Pyrite and the origin of life». Nature. 344 (6265): 387. Bibcode:1990Natur.344..387P. doi:10.1038/344387a0. S2CID 4322774.
- ^ a b Keller, Markus A.; Turchyn, Alexandra V.; Ralser, Markus (25 March 2014). «Non-enzymatic glycolysis and pentose phosphate pathway-like reactions in a plausible Archean ocean». Molecular Systems Biology. 10 (725): 725. doi:10.1002/msb.20145228. PMC 4023395. PMID 24771084.
- ^ Huber, Claudia; Wächtershäuser, Günter (31 July 1998). «Peptides by Activation of Amino Acids with CO on (Ni,Fe)S Surfaces: Implications for the Origin of Life». Science. 281 (5377): 670–672. Bibcode:1998Sci…281..670H. doi:10.1126/science.281.5377.670. PMID 9685253.
- ^ Adamala, Katarzyna; Szostak, Jack W. (29 November 2013). «Nonenzymatic Template-Directed RNA Synthesis Inside Model Protocells». Science. 342 (6162): 1098–1100. Bibcode:2013Sci…342.1098A. doi:10.1126/science.1241888. PMC 4104020. PMID 24288333.
- ^ Musser, George (23 September 2011). «How Life Arose on Earth, and How a Singularity Might Bring It Down». Observations (Blog). Archived from the original on 17 June 2015. Retrieved 17 June 2015.
- ^ Carroll, Sean (10 March 2010). «Free Energy and the Meaning of Life». Cosmic Variance (Blog). Discover. Archived from the original on 14 July 2015. Retrieved 17 June 2015.
- ^ England, Jeremy L. (28 September 2013). «Statistical physics of self-replication» (PDF). Journal of Chemical Physics. 139 (12): 121923. arXiv:1209.1179. Bibcode:2013JChPh.139l1923E. doi:10.1063/1.4818538. hdl:1721.1/90392. PMID 24089735. S2CID 478964. Archived (PDF) from the original on 4 June 2015.
- ^ Fox, Ronald F. (December 1993). «Review of Stuart Kauffman, The Origins of Order: Self-Organization and Selection in Evolution». Biophysical Journal. 65 (6): 2698–2699. Bibcode:1993BpJ….65.2698F. doi:10.1016/s0006-3495(93)81321-3. PMC 1226010.
- ^ Orgel, Leslie E. (7 November 2000). «Self-organizing biochemical cycles». PNAS. 97 (23): 12503–12507. Bibcode:2000PNAS…9712503O. doi:10.1073/pnas.220406697. PMC 18793. PMID 11058157.
- ^ Chandru, Kuhan; Gilbert, Alexis; Butch, Christopher; Aono, Masashi; Cleaves, Henderson James II (21 July 2016). «The Abiotic Chemistry of Thiolated Acetate Derivatives and the Origin of Life». Scientific Reports. 6 (29883): 29883. Bibcode:2016NatSR…629883C. doi:10.1038/srep29883. PMC 4956751. PMID 27443234.
- ^ Vallee, Yannick; Shalayel, Ibrahim; Ly, Kieu-Dung; Rao, K. V. Raghavendra; Paëpe, Gael De; Märker, Katharina; Milet, Anne (8 November 2017). «At the very beginning of life on Earth: the thiol-rich peptide (TRP) world hypothesis». The International Journal of Developmental Biology. 61 (8–9): 471–478. doi:10.1387/ijdb.170028yv. PMID 29139533.
- ^ a b Mulkidjanian, Armen Y. (24 August 2009). «On the origin of life in the zinc world: 1. Photosynthesizing, porous edifices built of hydrothermally precipitated zinc sulfide as cradles of life on Earth». Biology Direct. 4: 26. doi:10.1186/1745-6150-4-26. PMC 3152778. PMID 19703272.
- ^ Wächtershäuser, Günter (December 1988). «Before Enzymes and Templates: Theory of Surface Metabolism». Microbiological Reviews. 52 (4): 452–484. doi:10.1128/MMBR.52.4.452-484.1988. PMC 373159. PMID 3070320.
- ^ Mulkidjanian, Armen Y.; Galperin, Michael Y. (24 August 2009). «On the origin of life in the zinc world. 2. Validation of the hypothesis on the photosynthesizing zinc sulfide edifices as cradles of life on Earth». Biology Direct. 4: 27. doi:10.1186/1745-6150-4-27. PMC 2749021. PMID 19703275.
- ^ Macallum, A. B. (1 April 1926). «The Paleochemistry of the body fluids and tissues». Physiological Reviews. 6 (2): 316–357. doi:10.1152/physrev.1926.6.2.316.
- ^ Mulkidjanian, Armen Y.; Bychkov, Andrew Yu.; Dibrova, Daria V.; et al. (3 April 2012). «Origin of first cells at terrestrial, anoxic geothermal fields». PNAS. 109 (14): E821–E830. Bibcode:2012PNAS..109E.821M. doi:10.1073/pnas.1117774109. PMC 3325685. PMID 22331915.
- ^ See also Lankenau 2011, pp. 225–286, interconnecting the «Two RNA worlds» concept, and Davidovich, Chen; Belousoff, Matthew; Bashan, Anat; Yonath, Ada (September 2009). «The evolving ribosome: from non-coded peptide bond formation to sophisticated translation machinery». Research in Microbiology. 160 (7): 487–492. doi:10.1016/j.resmic.2009.07.004. PMID 19619641.
- ^ a b Plasson, Raphaël; Kondepudi, Dilip K.; Bersini, Hugues; et al. (August 2007). «Emergence of homochirality in far-from-equilibrium systems: Mechanisms and role in prebiotic chemistry». Chirality. 19 (8): 589–600. doi:10.1002/chir.20440. PMID 17559107. «Special Issue: Proceedings from the Eighteenth International Symposium on Chirality (ISCD-18), Busan, Korea, 2006»
- ^ Chaichian, Rojas & Tureanu 2014, pp. 353–364
- ^ Jafarpour, Farshid; Biancalani, Tommaso; Goldenfeld, Nigel (2017). «Noise-induced symmetry breaking far from equilibrium and the emergence of biological homochirality» (PDF). Physical Review E. 95 (3): 032407. Bibcode:2017PhRvE..95c2407J. doi:10.1103/PhysRevE.95.032407. PMID 28415353.
- ^ Jafarpour, Farshid; Biancalani, Tommaso; Goldenfeld, Nigel (2015). «Noise-induced mechanism for biological homochirality of early life self-replicators». Physical Review Letters. 115 (15): 158101. arXiv:1507.00044. Bibcode:2015PhRvL.115o8101J. doi:10.1103/PhysRevLett.115.158101. PMID 26550754. S2CID 9775791.
- ^ Frank, F.C. (1953). «On spontaneous asymmetric synthesis». Biochimica et Biophysica Acta. 11 (4): 459–463. doi:10.1016/0006-3002(53)90082-1. PMID 13105666.
- ^ Clark, Stuart (July–August 1999). «Polarized Starlight and the Handedness of Life». American Scientist. 87 (4): 336. Bibcode:1999AmSci..87..336C. doi:10.1511/1999.4.336.
- ^ Shibata, Takanori; Morioka, Hiroshi; Hayase, Tadakatsu; et al. (17 January 1996). «Highly Enantioselective Catalytic Asymmetric Automultiplication of Chiral Pyrimidyl Alcohol». Journal of the American Chemical Society. 118 (2): 471–472. doi:10.1021/ja953066g.
- ^ Soai, Kenso; Sato, Itaru; Shibata, Takanori (2001). «Asymmetric autocatalysis and the origin of chiral homogeneity in organic compounds». The Chemical Record. 1 (4): 321–332. doi:10.1002/tcr.1017. PMID 11893072.
- ^ Hazen 2005, p. 184
- ^ Meierhenrich, Uwe (2008). Amino acids and the asymmetry of life caught in the act of formation. Berlin: Springer. pp. 76–79. ISBN 978-3540768869.
- ^ Mullen, Leslie (5 September 2005). «Building Life from Star-Stuff». Astrobiology Magazine. Archived from the original on 14 July 2015.
General and cited sources[edit]
- Altermann, Wladyslaw (2009). «Introduction: A Roadmap to Fata Morgana?». In Seckbach, Joseph; Walsh, Maud (eds.). From Fossils to Astrobiology: Records of Life on Earth and the Search for Extraterrestrial Biosignatures. Cellular Origin, Life in Extreme Habitats and Astrobiology. Vol. 12. Dordrecht, the Netherlands; London: Springer. ISBN 978-1-4020-8836-0.
- Bada, Jeffrey L.; Lazcano, Antonio (2009). «The Origin of Life». In Ruse, Michael; Travis, Joseph (eds.). Evolution: The First Four Billion Years. Foreword by Edward O. Wilson. Cambridge: Belknap Press of Harvard University Press. ISBN 978-0-674-03175-3. OCLC 225874308.
- Barton, Nicholas H.; Briggs, Derek E.G.; Eisen, Jonathan A.; et al. (2007). Evolution. Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press. ISBN 978-0-87969-684-9. OCLC 86090399.
- Bastian, H. Charlton (1871). The Modes of Origin of Lowest Organisms. London; New York: Macmillan and Company. OCLC 42959303. Retrieved 6 June 2015.
- Bernal, J. D. (1951). The Physical Basis of Life. London: Routledge & Kegan Paul.
- Bernal, J. D. (1960). «The Problem of Stages in Biopoesis». In Florkin, M. (ed.). Aspects of the Origin of Life. International Series of Monographs on Pure and Applied Biology. Oxford, UK; New York: Pergamon Press. ISBN 978-1-4831-3587-8.
- Bernal, J. D. (1967) [Reprinted work by A.I. Oparin originally published 1924; Moscow: The Moscow Worker]. The Origin of Life. The Weidenfeld and Nicolson Natural History. Translation of Oparin by Ann Synge. London: Weidenfeld & Nicolson.
- Bock, Gregory R.; Goode, Jamie A., eds. (1996). Evolution of Hydrothermal Ecosystems on Earth (and Mars?). Ciba Foundation Symposium. Vol. 202. Chichester, UK; New York: John Wiley & Sons. ISBN 978-0-471-96509-1.
- Bondeson, Jan (1999). The Feejee Mermaid and Other Essays in Natural and Unnatural History. Ithaca, NY: Cornell University Press. ISBN 978-0-8014-3609-3.
- Bryson, Bill (2004). A Short History of Nearly Everything. London: Black Swan. ISBN 978-0-552-99704-1. OCLC 55589795.
- Calvin, Melvin (1969). Chemical Evolution: Molecular Evolution Towards the Origin of Living Systems on the Earth and Elsewhere. Oxford, UK: Clarendon Press. ISBN 978-0-19-855342-7. OCLC 25220.
- Chaichian, Masud; Rojas, Hugo Perez; Tureanu, Anca (2014). «Physics and Life». Basic Concepts in Physics: From the Cosmos to Quarks. Undergraduate Lecture Notes in Physics. Berlin; Heidelberg: Springer Berlin Heidelberg. doi:10.1007/978-3-642-19598-3_12. ISBN 978-3-642-19597-6. OCLC 900189038. S2CID 115247432.
- Chang, Thomas Ming Swi (2007). Artificial Cells: Biotechnology, Nanomedicine, Regenerative Medicine, Blood Substitutes, Bioencapsulation, and Cell/Stem Cell Therapy. Regenerative Medicine, Artificial Cells and Nanomedicine. Vol. 1. Hackensack, New Jersey: World Scientific. ISBN 978-981-270-576-1. OCLC 173522612.
- Dalrymple, G. Brent (2001). «The age of the Earth in the twentieth century: a problem (mostly) solved». In Lewis, C.L.E.; Knell, S.J. (eds.). The Age of the Earth: from 4004 BC to AD 2002. Geological Society of London, Special Publications. Geological Society Special Publication. Vol. 190. London: Geological Society of London. pp. 205–221. Bibcode:2001GSLSP.190..205D. doi:10.1144/gsl.sp.2001.190.01.14. ISBN 978-1-86239-093-5. OCLC 48570033. S2CID 130092094.
- Darwin, Charles (1887). Darwin, Francis (ed.). The Life and Letters of Charles Darwin, Including an Autobiographical Chapter. Vol. 3 (3rd ed.). London: John Murray. OCLC 834491774.
- Davies, Paul (1999). The Fifth Miracle: The Search for the Origin of Life. London: Penguin Books. ISBN 978-0-14-028226-9.
- Dawkins, Richard (1996). The Blind Watchmaker (Reissue with a new introduction ed.). New York: W.W. Norton & Company. ISBN 978-0-393-31570-7. OCLC 35648431.
- Dobell, Clifford (1960) [Originally published 1932; New York: Harcourt, Brace & Company]. Antony van Leeuwenhoek and His ‘Little Animals’. New York: Dover Publications.
- Dyson, Freeman (1999). Origins of Life (Revised ed.). Cambridge, UK; New York: Cambridge University Press. ISBN 978-0-521-62668-2.
- Fesenkov, V. G. (1959). «Some Considerations about the Primaeval State of the Earth». In Oparin, A.I.; et al. (eds.). The Origin of Life on the Earth. I.U.B. Symposium Series. Vol. 1. Edited for the International Union of Biochemistry by Frank Clark and R.L.M. Synge (English-French-German ed.). London; New York: Pergamon Press. ISBN 978-1-4832-2240-0. International Symposium on the Origin of Life on the Earth (held at Moscow, 19–24 August 1957)
- Hazen, Robert M. (2005). Genesis: The Scientific Quest for Life’s Origin. Washington, DC: Joseph Henry Press. ISBN 978-0-309-09432-0. OCLC 60321860.
- Huxley, Thomas Henry (1968) [1897]. «VIII Biogenesis and Abiogenesis [1870]». Discourses, Biological and Geological. Collected Essays. Vol. VIII (Reprint ed.). New York: Greenwood Press. OCLC 476737627.
- Klyce, Brig (22 January 2001). Kingsley, Stuart A.; Bhathal, Ragbir (eds.). Panspermia Asks New Questions. The Search for Extraterrestrial Intelligence (SETI) in the Optical Spectrum III. Vol. 4273. Bellingham, WA: SPIE. doi:10.1117/12.435366. ISBN 0-8194-3951-7. Retrieved 9 June 2015. Proceedings of the SPIE held at San Jose, California, 22–24 January 2001
- Lane, Nick (2009). Life Ascending: The 10 Great Inventions of Evolution (1st American ed.). New York: W.W. Norton & Company. ISBN 978-0-393-06596-1. OCLC 286488326.
- Lankenau, Dirk-Henner (2011). «Two RNA Worlds: Toward the Origin of Replication, Genes, Recombination and Repair». In Egel, Richard; Lankenau, Dirk-Henner; Mulkidjanian, Armen Y. (eds.). Origins of Life: The Primal Self-Organization. Heidelberg: Springer. doi:10.1007/978-3-642-21625-1. ISBN 978-3-642-21624-4. OCLC 733245537.
- Lennox, James G. (2001). Aristotle’s Philosophy of Biology: Studies in the Origins of Life Science. Cambridge Studies in Philosophy and Biology. Cambridge, UK; New York: Cambridge University Press. ISBN 978-0-521-65976-5.
- Michod, Richard E. (1999). Darwinian Dynamics: Evolutionary Transitions in Fitness and Individuality. Princeton, NJ: Princeton University Press. ISBN 978-0-691-02699-2. OCLC 38948118.
- Oparin, A.I. (1953) [Originally published 1938; New York: The Macmillan Company]. The Origin of Life. Translation and new introduction by Sergius Morgulis (2nd ed.). Mineola, NY: Dover Publications. ISBN 978-0-486-49522-4.
- Raven, Peter H.; Johnson, George B. (2002). Biology (6th ed.). Boston: McGraw-Hill. ISBN 978-0-07-112261-0. OCLC 45806501.
- Ross, Alexander (1652). Arcana Microcosmi. Vol. II. London. OCLC 614453394.
- Sheldon, Robert B. (22 September 2005). «Historical Development of the Distinction between Bio- and Abiogenesis» (PDF). In Hoover, Richard B.; Levin, Gilbert V.; Rozanov, Alexei Y.; Gladstone, G. Randall (eds.). Astrobiology and Planetary Missions. Astrobiology and Planetary Missions. Vol. 5906. Bellingham, WA: SPIE. pp. 59061I. doi:10.1117/12.663480. ISBN 978-0-8194-5911-4. Proceedings of the SPIE held at San Diego, California, 31 July–2 August 2005
- Tyndall, John (1905) [Originally published 1871; London; New York: Longmans, Green & Co.; D. Appleton and Company]. Fragments of Science. Vol. 2 (6th ed.). New York: P.F. Collier & Sons. OCLC 726998155.
- Voet, Donald; Voet, Judith G. (2004). Biochemistry. Vol. 1 (3rd ed.). New York: John Wiley & Sons. ISBN 978-0-471-19350-0.
- Yarus, Michael (2010). Life from an RNA World: The Ancestor Within. Cambridge, Massachusetts: Harvard University Press. p. 287. ISBN 978-0-674-05075-4.
External links[edit]
- Making headway with the mysteries of life’s origins – Adam Mann (PNAS; 14 April 2021)
- How life began on Earth – Marcia Malory (Earth Facts; 2015)
- The Origins of Life – Richard Dawkins et al. (BBC Radio; 2004)
- Life in the Universe – Essay by Stephen Hawking (1996)
«Origin of life» redirects here. For non-scientific views on the origins of life, see Creation myth.
Stages in the origin of life range from the well-understood, such as the habitable Earth and the abiotic synthesis of simple molecules, to the largely unknown, like the derivation of the last universal common ancestor (LUCA) with its complex molecular functionalities.[1]
In biology, abiogenesis (from a- ‘not’ + Greek bios ‘life’ + genesis ‘origin’) or the origin of life is the natural process by which life has arisen from non-living matter, such as simple organic compounds. The prevailing scientific hypothesis is that the transition from non-living to living entities on Earth was not a single event, but an evolutionary process (i.e., a process of gradually increasing complexity) that involved the formation of a habitable planet, the prebiotic synthesis of organic molecules, molecular self-replication, self-assembly, autocatalysis, and the emergence of cell membranes. Many proposals have been made for different stages of the process.
The study of abiogenesis aims to determine how pre-life chemical reactions gave rise to life under conditions strikingly different from those on Earth today. It primarily uses tools from biology and chemistry, with more recent approaches attempting a synthesis of many sciences. Life functions through the specialized chemistry of carbon and water, and builds largely upon four key families of chemicals: lipids for cell membranes, carbohydrates such as sugars, amino acids for protein metabolism, and nucleic acids DNA and RNA for the mechanisms of heredity. Any successful theory of abiogenesis must explain the origins and interactions of these classes of molecules. Many approaches to abiogenesis investigate how self-replicating molecules, or their components, came into existence. Researchers generally think that current life descends from an RNA world, although other self-replicating molecules may have preceded RNA.
The classic 1952 Miller–Urey experiment demonstrated that most amino acids, the chemical constituents of proteins, can be synthesized from inorganic compounds under conditions intended to replicate those of the early Earth. External sources of energy may have triggered these reactions, including lightning, radiation, atmospheric entries of micro-meteorites and implosion of bubbles in sea and ocean waves. Other approaches («metabolism-first» hypotheses) focus on understanding how catalysis in chemical systems on the early Earth might have provided the precursor molecules necessary for self-replication.
A genomics approach has sought to characterise the last universal common ancestor (LUCA) of modern organisms by identifying the genes shared by Archaea and Bacteria, members of the two major branches of life (where the Eukaryotes belong to the archaean branch in the two-domain system). 355 genes appear to be common to all life; their nature implies that the LUCA was anaerobic with the Wood–Ljungdahl pathway, deriving energy by chemiosmosis, and maintaining its hereditary material with DNA, the genetic code, and ribosomes. Although the LUCA lived over 4 billion years ago (4 Gya), researchers do not believe it was the first form of life. Earlier cells might have had a leaky membrane and been powered by a naturally-occurring proton gradient near a deep-sea white smoker hydrothermal vent.
Earth remains the only place in the universe known to harbor life, and fossil evidence from the Earth informs most studies of abiogenesis. The Earth was formed 4.54 Gya; the earliest undisputed evidence of life on Earth dates from at least 3.5 Gya. Fossil micro-organisms appear to have lived within hydrothermal vent precipitates dated 3.77 to 4.28 Gya from Quebec, soon after ocean formation 4.4 Gya during the Hadean.
Overview[edit]
NASA’s 2015 strategy for astrobiology aimed to solve the puzzle of the origin of life – how a fully-functioning living system could emerge from non-living components – through research on the prebiotic origin of life’s chemicals, both in space and on planets, as well as the functioning of early biomolecules to catalyse reactions and support inheritance.[2]
Life consists of reproduction with (heritable) variations.[3] NASA defines life as «a self-sustaining chemical system capable of Darwinian [i.e., biological] evolution.»[4] Such a system is complex; the last universal common ancestor (LUCA), presumably a single-celled organism which lived some 4 billion years ago, already had hundreds of genes encoded in the DNA genetic code that is universal today. That in turn implies a suite of cellular machinery including messenger RNA, transfer RNAs, and ribosomes to translate the code into proteins. Those proteins included enzymes to operate its anaerobic respiration via the Wood–Ljungdahl metabolic pathway, and a DNA polymerase to replicate its genetic material.[5][6]
The challenge for abiogenesis (origin of life)[7][8][9] researchers is to explain how such a complex and tightly-interlinked system could develop by evolutionary steps, as at first sight all its parts are necessary to enable it to function. For example, a cell, whether the LUCA or in a modern organism, copies its DNA with the DNA polymerase enzyme, which is in turn produced by translating the DNA polymerase gene in the DNA. Neither the enzyme nor the DNA can be produced without the other.[10] The evolutionary process could have involved molecular self-replication, self-assembly such as of cell membranes, and autocatalysis.[5][6][11]
The precursors to the development of a living cell like the LUCA are clear enough, if disputed in their details: a habitable world is formed with a supply of minerals and liquid water. Prebiotic synthesis creates a range of simple organic compounds, which are assembled into polymers such as proteins and RNA. The process after the LUCA, too, is readily understood: biological evolution caused the development of a wide range of species with varied forms and biochemical capabilities. The derivation of living things such as the LUCA from simple components, however, is far from understood.[1]
Although Earth remains the only place where life is known,[12][13] the science of astrobiology seeks evidence of life on other planets. The 2015 NASA strategy on the origin of life aimed to solve the puzzle by identifying interactions, intermediary structures and functions, energy sources, and environmental factors that contributed to the diversity, selection, and replication of evolvable macromolecular systems,[2] and mapping the chemical landscape of potential primordial informational polymers. The advent of polymers that could replicate, store genetic information, and exhibit properties subject to selection was, it suggested, most likely a critical step in the emergence of prebiotic chemical evolution.[2] Those polymers derived, in turn, from simple organic compounds such as nucleobases, amino acids and sugars that could have been formed by reactions in the environment.[14][8][15][16] A successful theory of the origin of life must explain how all these chemicals came into being.[17]
Conceptual history until the 1960s[edit]
The Miller–Urey experiment was a synthesis of small organic molecules in a mixture of simple gases in a thermal gradient created by heating (right) and cooling (left) the mixture at the same time, with electrical discharges.
Spontaneous generation[edit]
One ancient view of the origin of life, from Aristotle until the 19th century, is of spontaneous generation.[18] This theory held that «lower» animals were generated by decaying organic substances, and that life arose by chance.[19][20] This was questioned from the 17th century, in works like Thomas Browne’s Pseudodoxia Epidemica.[21][22] In 1665, Robert Hooke published the first drawings of a microorganism. In 1676, Antonie van Leeuwenhoek drew and described microorganisms, probably protozoa and bacteria.[23] Van Leeuwenhoek disagreed with spontaneous generation, and by the 1680s convinced himself, using experiments ranging from sealed and open meat incubation and the close study of insect reproduction, that the theory was incorrect.[24] In 1668 Francesco Redi showed that no maggots appeared in meat when flies were prevented from laying eggs.[25] By the middle of the 19th century, spontaneous generation was considered disproven.[26][27]
Panspermia[edit]
Another ancient idea dating back to Anaxagoras in the 5th century BC is panspermia,[28] the idea that life exists throughout the universe, distributed by meteoroids, asteroids, comets[29] and planetoids.[30] It does not attempt to explain how life originated in itself, but shifts the origin of life on Earth to another heavenly body. The advantage is that life is not required to have formed on each planet it occurs on, but rather in a more limited set of locations (potentially even a single location), and then spread about the galaxy to other star systems via cometary or meteorite impact.[31]
«A warm little pond»: primordial soup[edit]
The idea that life originated from non-living matter in slow stages appeared in Herbert Spencer’s 1864–1867 book Principles of Biology, and in William Turner Thiselton-Dyer’s 1879 paper «On spontaneous generation and evolution». On 1 February 1871 Charles Darwin wrote about these publications to Joseph Hooker, and set out his own speculation, suggesting that the original spark of life may have begun in a «warm little pond, with all sorts of ammonia and phosphoric salts, light, heat, electricity, &c., present, that a proteine compound was chemically formed ready to undergo still more complex changes.» Darwin went on to explain that «at the present day such matter would be instantly devoured or absorbed, which would not have been the case before living creatures were formed.»[32][33][34]
Alexander Oparin in 1924 and J. B. S. Haldane in 1929 proposed that the first molecules constituting the earliest cells slowly self-organized from a primordial soup, and this theory is called the Oparin–Haldane hypothesis.[35][36] Haldane suggested that the Earth’s prebiotic oceans consisted of a «hot dilute soup» in which organic compounds could have formed.[20][37] J. D. Bernal showed that such mechanisms could form most of the necessary molecules for life from inorganic precursors.[38] In 1967, he suggested three «stages»: the origin of biological monomers; the origin of biological polymers; and the evolution from molecules to cells.[39][40]
Miller–Urey experiment[edit]
In 1952, Stanley Miller and Harold Urey carried out a chemical experiment to demonstrate how organic molecules could have formed spontaneously from inorganic precursors under prebiotic conditions like those posited by the Oparin–Haldane hypothesis. It used a highly reducing (lacking oxygen) mixture of gases—methane, ammonia, and hydrogen, as well as water vapor—to form simple organic monomers such as amino acids.[41][42] Bernal said of the Miller–Urey experiment that «it is not enough to explain the formation of such molecules, what is necessary, is a physical-chemical explanation of the origins of these molecules that suggests the presence of suitable sources and sinks for free energy.»[43] However, current scientific consensus describes the primitive atmosphere as weakly reducing or neutral,[44][45] diminishing the amount and variety of amino acids that could be produced. The addition of iron and carbonate minerals, present in early oceans, however produces a diverse array of amino acids.[44] Later work has focused on two other potential reducing environments: outer space and deep-sea hydrothermal vents.[46][47][48]
Producing a habitable Earth[edit]
Early universe with first stars[edit]
Soon after the Big Bang, which occurred roughly 14 Gya, the only chemical elements present in the universe were hydrogen, helium, and lithium, the three lightest atoms in the periodic table. These elements gradually came together to form stars. These early stars were massive and short-lived, producing all the heavier elements through stellar nucleosynthesis. Carbon, currently the fourth most abundant chemical element in the universe (after hydrogen, helium and oxygen), was formed mainly in white dwarf stars, particularly those bigger than twice the mass of the sun.[49] As these stars reached the end of their lifecycles, they ejected these heavier elements, among them carbon and oxygen, throughout the universe. These heavier elements allowed for the formation of new objects, including rocky planets and other bodies.[50] According to the nebular hypothesis, the formation and evolution of the Solar System began 4.6 Gya with the gravitational collapse of a small part of a giant molecular cloud. Most of the collapsing mass collected in the center, forming the Sun, while the rest flattened into a protoplanetary disk out of which the planets, moons, asteroids, and other small Solar System bodies formed.[51]
Emergence of Earth[edit]
The Earth was formed 4.54 Gya.[52][53] The Hadean Earth (from its formation until 4 Gya) was at first inhospitable to any living organisms. During its formation, the Earth lost a significant part of its initial mass, and consequentially lacked the gravity to hold molecular hydrogen and the bulk of the original inert gases.[54] The atmosphere consisted largely of water vapor, nitrogen, and carbon dioxide, with smaller amounts of carbon monoxide, hydrogen, and sulfur compounds.[55] The solution of carbon dioxide in water is thought to have made the seas slightly acidic, with a pH of about 5.5.[56] The Hadean atmosphere has been characterized as a «gigantic, productive outdoor chemical laboratory,»[57] similar to volcanic gases today which still support some abiotic chemistry.[57]
Oceans may have appeared as soon as 200 million years after the Earth formed, in a near-boiling (100 C) reducing environment, as the pH of 5.8 rose rapidly towards neutral.[58] This scenario has found support from the dating of 4.404 Gya zircon crystals from metamorphosed quartzite of Mount Narryer in Western Australia.[59] Despite the likely increased volcanism, the Earth may have been a water world between 4.4 and 4.3 Gya, with little if any continental crust, a turbulent atmosphere, and a hydrosphere subject to intense ultraviolet light from a T Tauri stage Sun, from cosmic radiation, and from continued asteroid and comet impacts.[60]
The Late Heavy Bombardment hypothesis posits that the Hadean environment between 4.28[61] and 3.8 Gya was highly hazardous to life. Following the Nice model, changes in the orbits of the giant planets may have bombarded the Earth with asteroids and comets that pockmarked the Moon and inner planets.[62] Frequent collisions would have made photosynthesis unviable.[57][63][64][65] The periods between such devastating events give time windows for the possible origin of life in early environments. If the deep marine hydrothermal setting was the site for the origin of life, then abiogenesis could have happened as early as 4.0-4.2 Gya. If the site was at the surface of the Earth, abiogenesis could have occurred only between 3.7 and 4.0 Gya.[66] However, new lunar surveys and samples have led scientists, including an architect of the Nice model, to deemphasize the significance of the Late Heavy Bombardment.[67]
If life evolved in the ocean at depths of more than ten meters, it would have been shielded both from late impacts and the then high levels of ultraviolet radiation from the sun. Geothermically heated oceanic crust could have yielded far more organic compounds through deep hydrothermal vents than the Miller–Urey experiments indicated.[68] The available energy is maximized at 100–150 °C, the temperatures at which hyperthermophilic bacteria and thermoacidophilic archaea live. These modern organisms may be among the closest surviving relatives of the LUCA.[69]
Earliest evidence of life[edit]
Life existed on Earth more than 3.5 Gya,[70][71][72] during the Eoarchean when sufficient crust had solidified following the molten Hadean.[73][74][75] The earliest physical evidence of life so far found consists of microfossils in the Nuvvuagittuq Greenstone Belt of Northern Quebec, in banded iron formation rocks at least 3.77 and possibly 4.28 Gya. The micro-organisms lived within hydrothermal vent precipitates, soon after the 4.4 Gya formation of oceans during the Hadean. The microbes resembled modern hydrothermal vent bacteria, supporting the view that abiogenesis began in such an environment.[61]
Biogenic graphite has been found in 3.7 Gya metasedimentary rocks from southwestern Greenland[76] and in microbial mat fossils from 3.49 Gya Western Australian sandstone.[77] Evidence of early life in rocks from Akilia Island, near the Isua supracrustal belt in southwestern Greenland, dating to 3.7 Gya, have shown biogenic carbon isotopes.[78] In other parts of the Isua supracrustal belt, graphite inclusions trapped within garnet crystals are connected to the other elements of life: oxygen, nitrogen, and possibly phosphorus in the form of phosphate, providing further evidence for life 3.7 Gya.[79] In the Pilbara region of Western Australia, compelling evidence of early life was found in pyrite-bearing sandstone in a fossilized beach, with rounded tubular cells that oxidized sulfur by photosynthesis in the absence of oxygen.[80][81] Zircons from Western Australia imply that life existed on Earth at least 4.1 Gya.[82]
The Pilbara region of Western Australia contains the Dresser Formation with rocks 3.48 Gya, including layered structures called stromatolites. Their modern counterparts are created by photosynthetic micro-organisms including cyanobacteria.[83] These lie within undeformed hydrothermal-sedimentary strata; their texture indicates a biogenic origin. Parts of the Dresser formation preserve hot springs on land, but other regions seem to have been shallow seas.[84]
Producing molecules: prebiotic synthesis[edit]
All chemical elements except for hydrogen and helium derive from stellar nucleosynthesis. The basic chemical ingredients of life – the carbon-hydrogen molecule (CH), the carbon-hydrogen positive ion (CH+) and the carbon ion (C+) – were produced by ultraviolet light from stars.[85] Complex molecules, including organic molecules, form naturally both in space and on planets.[86] Organic molecules on the early Earth could have had either terrestrial origins, with organic molecule synthesis driven by impact shocks or by other energy sources, such as ultraviolet light, redox coupling, or electrical discharges; or extraterrestrial origins (pseudo-panspermia), with organic molecules formed in interstellar dust clouds raining down on to the planet.[87][88]
[edit]
An organic compound is a chemical whose molecules contain carbon. Carbon is abundant in the Sun, stars, comets, and in the atmospheres of most planets.[89] Organic compounds are relatively common in space, formed by «factories of complex molecular synthesis» which occur in molecular clouds and circumstellar envelopes, and chemically evolve after reactions are initiated mostly by ionizing radiation.[86][90][91] Purine and pyrimidine nucleobases including guanine, adenine, cytosine, uracil and thymine have been found in meteorites. These could have provided the materials for DNA and RNA to form on the early Earth.[92] The amino acid glycine was found in material ejected from comet Wild 2; it had earlier been detected in meteorites.[93] Comets are encrusted with dark material, thought to be a tar-like organic substance formed from simple carbon compounds under ionizing radiation. A rain of material from comets could have brought such complex organic molecules to Earth.[94][95][57] It is estimated that during the Late Heavy Bombardment, meteorites may have delivered up to five million tons of organic prebiotic elements to Earth per year.[57]
PAH world hypothesis[edit]
Polycyclic aromatic hydrocarbons (PAH) are the most common and abundant polyatomic molecules in the observable universe, and are a major store of carbon.[89][96][97][98] They seem to have formed shortly after the Big Bang,[99][97][98] and are associated with new stars and exoplanets.[89] They are a likely constituent of Earth’s primordial sea.[99][97][98] PAHs have been detected in nebulae,[100] and in the interstellar medium, in comets, and in meteorites.[89]
The PAH world hypothesis posits PAHs as precursors to the RNA world.[101] A star, HH 46-IR, resembling the sun early in its life, is surrounded by a disk of material which contains molecules including cyanide compounds, hydrocarbons, and carbon monoxide. PAHs in the interstellar medium can be transformed through hydrogenation, oxygenation, and hydroxylation to more complex organic compounds used in living cells.[102]
Nucleobases[edit]
The majority of organic compounds introduced on Earth by interstellar dust particles have helped to form complex molecules, thanks to their peculiar surface-catalytic activities.[103][104] Studies of the 12C/13C isotopic ratios of organic compounds in the Murchison meteorite suggest that the RNA component uracil and related molecules, including xanthine, were formed extraterrestrially.[105] NASA studies of meteorites suggest that all four DNA nucleobases (adenine, guanine and related organic molecules) have been formed in outer space.[103][106][107] The cosmic dust permeating the universe contains complex organics («amorphous organic solids with a mixed aromatic–aliphatic structure») that could be created rapidly by stars.[108] Glycolaldehyde, a sugar molecule and RNA precursor, has been detected in regions of space including around protostars and on meteorites.[109][110]
Laboratory synthesis[edit]
As early as the 1860s, experiments demonstrated that biologically relevant molecules can be produced from interaction of simple carbon sources with abundant inorganic catalysts. The spontaneous formation of complex polymers from abiotically generated monomers under the conditions posited by the «soup» theory is not straightforward. Besides the necessary basic organic monomers, compounds that would have prohibited the formation of polymers were also formed in high concentration during the Miller–Urey and Joan Oró experiments.[111] Biology uses essentially 20 amino acids for its coded protein enzymes, representing a very small subset of the structurally possible products. Since life tends to use whatever is available, an explanation is needed for why the set used is so small.[112]
Sugars[edit]
The Breslow catalytic cycle for formaldehyde dimerization and C2-C6 sugar formation
Alexander Butlerov showed in 1861 that the formose reaction created sugars including tetroses, pentoses, and hexoses when formaldehyde is heated under basic conditions with divalent metal ions like calcium. R. Breslow proposed that the reaction was autocatalytic in 1959.[113]
Nucleobases[edit]
Nucleobases like guanine and adenine can be synthesized from simple carbon and nitrogen sources like hydrogen cyanide (HCN) and ammonia.[114] Formamide produces all four ribonucleotides when warmed with terrestrial minerals. Formamide is ubiquitous in the Universe, produced by the reaction of water and HCN. It can be concentrated by the evaporation of water.[115][116] HCN is poisonous only to aerobic organisms (eukaryotes and aerobic bacteria), which did not yet exist. It can play roles in other chemical processes such as the synthesis of the amino acid glycine.[57]
DNA and RNA components including uracil, cytosine and thymine can be synthesized under outer space conditions, using starting chemicals such as pyrimidine found in meteorites. Pyrimidine may have been formed in red giant stars or in interstellar dust and gas clouds.[117] All four RNA-bases may be synthesized from formamide in high-energy density events like extraterrestrial impacts.[118]
Other pathways for synthesizing bases from inorganic materials have been reported.[119] Freezing temperatures are advantageous for the synthesis of purines, due to the concentrating effect for key precursors such as hydrogen cyanide.[120] However, while adenine and guanine require freezing conditions for synthesis, cytosine and uracil may require boiling temperatures.[121] Seven different amino acids and eleven types of nucleobases formed in ice when ammonia and cyanide were left in a freezer for 25 years.[122][123] S-triazines (alternative nucleobases), pyrimidines including cytosine and uracil, and adenine can be synthesized by subjecting a urea solution to freeze-thaw cycles under a reductive atmosphere, with spark discharges as an energy source.[124] The explanation given for the unusual speed of these reactions at such a low temperature is eutectic freezing, which crowds impurities in microscopic pockets of liquid within the ice, causing the molecules to collide more often.[125]
Producing suitable vesicles[edit]
The lipid world theory postulates that the first self-replicating object was lipid-like.[126][127] Phospholipids form lipid bilayers in water while under agitation—the same structure as in cell membranes. These molecules were not present on early Earth, but other amphiphilic long-chain molecules also form membranes. These bodies may expand by insertion of additional lipids, and may spontaneously split into two offspring of similar size and composition. The main idea is that the molecular composition of the lipid bodies is a preliminary to information storage, and that evolution led to the appearance of polymers like RNA that store information. Studies on vesicles from amphiphiles that might have existed in the prebiotic world have so far been limited to systems of one or two types of amphiphiles.[128]
A lipid bilayer membrane could be composed of a huge number of combinations of arrangements of amphiphiles. The best of these would have favored the constitution of a hypercycle,[129][130] actually a positive feedback composed of two mutual catalysts represented by a membrane site and a specific compound trapped in the vesicle. Such site/compound pairs are transmissible to the daughter vesicles leading to the emergence of distinct lineages of vesicles, which would have allowed natural selection.[131]
A protocell is a self-organized, self-ordered, spherical collection of lipids proposed as a stepping-stone to the origin of life.[128] The theory of classical irreversible thermodynamics treats self-assembly under a generalized chemical potential within the framework of dissipative systems.[132][133][134]
A central question in evolution is how simple protocells first arose and differed in reproductive contribution to the following generation, thus driving the evolution of life. A functional protocell has (as of 2014) not yet been achieved in a laboratory setting.[135][136][137] Self-assembled vesicles are essential components of primitive cells.[128] The second law of thermodynamics requires that the universe move in a direction in which entropy increases, yet life is distinguished by its great degree of organization. Therefore, a boundary is needed to separate life processes from non-living matter.[138] Irene Chen and Jack W. Szostak suggest that elementary protocells can give rise to cellular behaviors including primitive forms of differential reproduction, competition, and energy storage.[136] Competition for membrane molecules would favor stabilized membranes, suggesting a selective advantage for the evolution of cross-linked fatty acids and even the phospholipids of today.[136] Such micro-encapsulation would allow for metabolism within the membrane and the exchange of small molecules, while retaining large biomolecules inside. Such a membrane is needed for a cell to create its own electrochemical gradient to store energy by pumping ions across the membrane.[139][140]
Producing biology[edit]
Energy and entropy[edit]
Life requires a loss of entropy, or disorder, when molecules organize themselves into living matter. The emergence of life and increased complexity does not contradict the second law of thermodynamics, which states that overall entropy never decreases, since a living organism creates order in some places (e.g. its living body) at the expense of an increase of entropy elsewhere (e.g. heat and waste production).[141][142][143]
Multiple sources of energy were available for chemical reactions on the early Earth. Heat from geothermal processes is a standard energy source for chemistry. Other examples include sunlight, lightning,[57] atmospheric entries of micro-meteorites,[144] and implosion of bubbles in sea and ocean waves.[145] This has been confirmed by experiments[146][147] and simulations.[148]
Unfavorable reactions can be driven by highly favorable ones, as in the case of iron-sulfur chemistry. For example, this was probably important for carbon fixation.[a] Carbon fixation by reaction of CO2 with H2S via iron-sulfur chemistry is favorable, and occurs at neutral pH and 100 °C. Iron-sulfur surfaces, which are abundant near hydrothermal vents, can drive the production of small amounts of amino acids and other biomolecules.[57]
Chemiosmosis[edit]
In 1961, Peter Mitchell proposed chemiosmosis as a cell’s primary system of energy conversion. The mechanism, now ubiquitous in living cells, powers energy conversion in micro-organisms and in the mitochondria of eukaryotes, making it a likely candidate for early life.[149][150] Mitochondria produce adenosine triphosphate (ATP), the energy currency of the cell used to drive cellular processes such as chemical syntheses. The mechanism of ATP synthesis involves a closed membrane in which the ATP synthase enzyme is embedded. The energy required to release strongly-bound ATP has its origin in protons that move across the membrane.[151] In modern cells, those proton movements are caused by the pumping of ions across the membrane, maintaining an electrochemical gradient. In the first organisms, the gradient could have been provided by the difference in chemical composition between the flow from a hydrothermal vent and the surrounding seawater,[140] or perhaps meteoric quinones that were conducive to the development of chemiosmotic energy across lipid membranes if at a terrestrial origin.[152]
The RNA world[edit]
The RNA world hypothesis describes an early Earth with self-replicating and catalytic RNA but no DNA or proteins.[153] Many researchers concur that an RNA world must have preceded the DNA-based life that now dominates.[154] However, RNA-based life may not have been the first to exist.[155][156] Another model echoes Darwin’s «warm little pond» with cycles of wetting and drying.[157]
RNA is central to the translation process. Small RNAs can catalyze all the chemical groups and information transfers required for life.[156][158] RNA both expresses and maintains genetic information in modern organisms; and the chemical components of RNA are easily synthesized under the conditions that approximated the early Earth, which were very different from those that prevail today. The structure of the ribozyme has been called the «smoking gun», with a central core of RNA and no amino acid side chains within 18 Å of the active site that catalyzes peptide bond formation.[159][155][160]
The concept of the RNA world was proposed in 1962 by Alexander Rich,[161] and the term was coined by Walter Gilbert in 1986.[156][162] There were initial difficulties in the explanation of the abiotic synthesis of the nucleotides cytosine and uracil.[163] Subsequent research has shown possible routes of synthesis; for example, formamide produces all four ribonucleotides and other biological molecules when warmed in the presence of various terrestrial minerals.[115][116]
The RNA world hypothesis proposes that undirected polymerisation led to the emergence of ribozymes, and in turn to an RNA replicase.
RNA replicase can function as both code and catalyst for further RNA replication, i.e. it can be autocatalytic. Jack Szostak has shown that certain catalytic RNAs can join smaller RNA sequences together, creating the potential for self-replication. The RNA replication systems, which include two ribozymes that catalyze each other’s synthesis, showed a doubling time of the product of about one hour, and were subject to natural selection under the experimental conditions.[164][165][155] If such conditions were present on early Earth, then natural selection would favor the proliferation of such autocatalytic sets, to which further functionalities could be added.[166][167][168] Self-assembly of RNA may occur spontaneously in hydrothermal vents.[169][170][171] A preliminary form of tRNA could have assembled into such a replicator molecule.[172]
Possible precursors to protein synthesis include the synthesis of short peptide cofactors or the self-catalysing duplication of RNA. It is likely that the ancestral ribosome was composed entirely of RNA, although some roles have since been taken over by proteins. Major remaining questions on this topic include identifying the selective force for the evolution of the ribosome and determining how the genetic code arose.[173]
Eugene Koonin has argued that «no compelling scenarios currently exist for the origin of replication and translation, the key processes that together comprise the core of biological systems and the apparent pre-requisite of biological evolution. The RNA World concept might offer the best chance for the resolution of this conundrum but so far cannot adequately account for the emergence of an efficient RNA replicase or the translation system.»[174]
Phylogeny and LUCA[edit]
Starting with the work of Carl Woese from 1977 onwards, genomics studies have placed the last universal common ancestor (LUCA) of all modern life-forms between Bacteria and a clade formed by Archaea and Eukaryota in the phylogenetic tree of life. It lived over 4 Gya.[175][176] A minority of studies have placed the LUCA in Bacteria, proposing that Archaea and Eukaryota are evolutionarily derived from within Eubacteria;[177] Thomas Cavalier-Smith suggested that the phenotypically diverse bacterial phylum Chloroflexota contained the LUCA.[178]
In 2016, a set of 355 genes likely present in the LUCA was identified. A total of 6.1 million prokaryotic genes from Bacteria and Archaea were sequenced, identifying 355 protein clusters from amongst 286,514 protein clusters that were probably common to the LUCA. The results suggest that the LUCA was anaerobic with a Wood–Ljungdahl pathway, nitrogen- and carbon-fixing, thermophilic. Its cofactors suggest dependence upon an environment rich in hydrogen, carbon dioxide, iron, and transition metals. Its genetic material was probably DNA, requiring the 4-nucleotide genetic code, messenger RNA, transfer RNAs, and ribosomes to translate the code into proteins such as enzymes. LUCA likely inhabited an anaerobic hydrothermal vent setting in a geochemically active environment. It was evidently already a complex organism, and must have had precursors; it was not the first living thing.[10][179] The physiology of LUCA has been in dispute.[180][181][182]
-
LUCA systems and environment[10]
Leslie Orgel argued that early translation machinery for the genetic code would be susceptible to error catastrophe. Geoffrey Hoffmann however showed that such machinery can be stable in function against «Orgel’s paradox».[183][184][185]
Suitable geological environments[edit]
Deep sea hydrothermal vents[edit]
Early micro-fossils may have come from a hot world of gases such as methane, ammonia, carbon dioxide and hydrogen sulphide, toxic to much current life.[186] Analysis of the tree of life places thermophilic and hyperthermophilic bacteria and archaea closest to the root, suggesting that life may have evolved in a hot environment.[187] The deep sea or alkaline hydrothermal vent theory posits that life began at submarine hydrothermal vents.[188][189] Martin and Russell have suggested «that life evolved in structured iron monosulphide precipitates in a seepage site hydrothermal mound at a redox, pH, and temperature gradient between sulphide-rich hydrothermal fluid and iron(II)-containing waters of the Hadean ocean floor. The naturally arising, three-dimensional compartmentation observed within fossilized seepage-site metal sulphide precipitates indicates that these inorganic compartments were the precursors of cell walls and membranes found in free-living prokaryotes. The known capability of FeS and NiS to catalyze the synthesis of the acetyl-methylsulphide from carbon monoxide and methylsulphide, constituents of hydrothermal fluid, indicates that pre-biotic syntheses occurred at the inner surfaces of these metal-sulphide-walled compartments».[190]
These form where hydrogen-rich fluids emerge from below the sea floor, as a result of serpentinization of ultra-mafic olivine with seawater and a pH interface with carbon dioxide-rich ocean water. The vents form a sustained chemical energy source derived from redox reactions, in which electron donors (molecular hydrogen) react with electron acceptors (carbon dioxide); see Iron–sulfur world theory. These are exothermic reactions.[188][b]
Early cell powered by external proton gradient near a deep-sea hydrothermal vent. As long as the membrane (or passive ion channels within it) is permeable to protons, the mechanism can function without ion pumps.[140]
Russell demonstrated that alkaline vents created an abiogenic proton motive force chemiosmotic gradient,[190] ideal for abiogenesis. Their microscopic compartments «provide a natural means of concentrating organic molecules,» composed of iron-sulfur minerals such as mackinawite, endowed these mineral cells with the catalytic properties envisaged by Günter Wächtershäuser.[191] This movement of ions across the membrane depends on a combination of two factors:
- Diffusion force caused by concentration gradient—all particles including ions tend to diffuse from higher concentration to lower.
- Electrostatic force caused by electrical potential gradient—cations like protons H+ tend to diffuse down the electrical potential, anions in the opposite direction.
These two gradients taken together can be expressed as an electrochemical gradient, providing energy for abiogenic synthesis. The proton motive force can be described as the measure of the potential energy stored as a combination of proton and voltage gradients across a membrane (differences in proton concentration and electrical potential).[140]
The surfaces of mineral particles inside deep-ocean hydrothermal vents have catalytic properties similar to those of enzymes and can create simple organic molecules, such as methanol (CH3OH), formic, acetic and pyruvic acids out of the dissolved CO2 in the water, if driven by an applied voltage or by reaction with H2 or H2S.[192][193]
The research reported by Martin in 2016 supports the thesis that life arose at hydrothermal vents,[194][195] that spontaneous chemistry in the Earth’s crust driven by rock–water interactions at disequilibrium thermodynamically underpinned life’s origin[196][197] and that the founding lineages of the archaea and bacteria were H2-dependent autotrophs that used CO2 as their terminal acceptor in energy metabolism.[198] Martin suggests, based upon this evidence, that the LUCA «may have depended heavily on the geothermal energy of the vent to survive».[199] Pores at deep sea hydrothermal vents are suggested to have been occupied
by membrane-bound compartments which promoted biochemical reactions.[200][201]
Hot springs[edit]
Mulkidjanian and co-authors think that marine environments did not provide the ionic balance and composition universally found in cells, or the ions required by essential proteins and ribozymes, especially with respect to high K+/Na+ ratio, Mn2+, Zn2+ and phosphate concentrations. They argue that the only environments that mimic the needed conditions on Earth are hot springs similar to ones at Kamchatka.[202] Mineral deposits in these environments under an anoxic atmosphere would have suitable pH (while current pools in an oxygenated atmosphere would not), contain precipitates of photocatalytic sulfide minerals that absorb harmful ultraviolet radiation, have wet-dry cycles that concentrate substrate solutions to concentrations amenable to spontaneous formation of biopolymers[203][204] created both by chemical reactions in the hydrothermal environment, and by exposure to UV light during transport from vents to adjacent pools that would promote the formation of biomolecules.[205] The hypothesized pre-biotic environments are similar to hydrothermal vents, with additional components that help explain peculiarities of the LUCA.[202][152]
A phylogenomic and geochemical analysis of proteins plausibly traced to the LUCA shows that the ionic composition of its intracellular fluid is identical at hot springs. The LUCA likely was dependent upon synthesized organic matter for its growth.[202] Experiments show that RNA-like polymers can be synthesized in multiple wet-dry cycles and exposure to UV light. These polymers became encapsulated in vesicles after condensation, which would not happen in saltwater conditions because of the high concentrations of ionic solutes.[206] A potential source of biomolecules at hot springs was transport by interplanetary dust particles, extraterrestrial projectiles, or atmospheric or geochemical synthesis. Hot spring fields could have been abundant at volcanic landmasses during the Hadean.[152]
Clay[edit]
The clay hypothesis was proposed by Graham Cairns-Smith in 1985.[207][208] It postulates that complex organic molecules arose gradually on pre-existing, non-organic replication surfaces of silicate crystals in contact with an aqueous solution. The clay mineral montmorillonite has been shown to catalyze the polymerization of RNA in aqueous solution from nucleotide monomers,[209] and the formation of membranes from lipids.[210] In 1998, Hyman Hartman proposed that «the first organisms were self-replicating iron-rich clays which fixed carbon dioxide into oxalic acid and other dicarboxylic acids. This system of replicating clays and their metabolic phenotype then evolved into the sulfide rich region of the hot spring acquiring the ability to fix nitrogen. Finally phosphate was incorporated into the evolving system which allowed the synthesis of nucleotides and phospholipids.»[211]
Iron–sulfur world[edit]
In the 1980s, Günter Wächtershäuser and Karl Popper postulated the Iron–sulfur world hypothesis for the evolution of pre-biotic chemical pathways. It traces today’s biochemistry to primordial reactions which synthesize organic building blocks from gases.[212][213] Wächtershäuser systems have a built-in source of energy: iron sulfides such as pyrite. The energy released by oxidising these metal sulfides can support synthesis of organic molecules. Such systems may have evolved into autocatalytic sets constituting self-replicating, metabolically active entities predating modern life forms.[214] Experiments with sulfides in an aqueous environment at 100 °C produced a small yield of dipeptides (0.4% to 12.4%) and a smaller yield of tripeptides (0.10%). However, under the same conditions, dipeptides were quickly broken down.[215]
Several models postulate a primitive metabolism, allowing RNA replication to emerge later. The centrality of the Krebs cycle (citric acid cycle) to energy production in aerobic organisms, and in drawing in carbon dioxide and hydrogen ions in biosynthesis of complex organic chemicals, suggests that it was one of the first parts of the metabolism to evolve.[191] Concordantly, geochemists Jack W. Szostak and Kate Adamala demonstrated that non-enzymatic RNA replication in primitive protocells is only possible in the presence of weak cation chelators like citric acid. This provides further evidence for the central role of citric acid in primordial metabolism.[216]
Russell has proposed that «the purpose of life is to hydrogenate carbon dioxide» (as part of a «metabolism-first,» rather than a «genetics-first,» scenario).[217][218][214] The physicist Jeremy England has argued from general thermodynamic considerations that life was inevitable.[219] An early version of this idea was Oparin’s 1924 proposal for self-replicating vesicles. In the 1980s and 1990s came Wächtershäuser’s iron–sulfur world theory and Christian de Duve’s thioester models. More abstract and theoretical arguments for metabolism without genes include Freeman Dyson’s mathematical model and Stuart Kauffman’s collectively autocatalytic sets in the 1980s. Kauffman’s work has been criticized for ignoring the role of energy in driving biochemical reactions in cells.[220]
A multistep biochemical pathway like the Krebs cycle did not just self-organize on the surface of a mineral; it must have been preceded by simpler pathways.[221] The Wood–Ljungdahl pathway is compatible with self-organization on a metal sulfide surface. Its key enzyme unit, carbon monoxide dehydrogenase/acetyl-CoA synthase, contains mixed nickel-iron-sulfur clusters in its reaction centers and catalyzes the formation of acetyl-CoA. However, prebiotic thiolated and thioester compounds are thermodynamically and kinetically unlikely to accumulate in the presumed prebiotic conditions of hydrothermal vents.[222] One possibility is that cysteine and homocysteine may have reacted with nitriles from the Stecker reaction, forming catalytic thiol-rich polypeptides.[223]
Zinc world[edit]
Armen Mulkidjanian’s zinc world (Zn-world) hypothesis extends Wächtershäuser’s pyrite hypothesis.[224][225] The Zn-world theory proposes that hydrothermal fluids rich in H2S interacting with cold primordial ocean (or Darwin’s «warm little pond») water precipitated metal sulfide particles. Oceanic hydrothermal systems have a zonal structure reflected in ancient volcanogenic massive sulfide ore deposits. They reach many kilometers in diameter and date back to the Archean. Most abundant are pyrite (FeS2), chalcopyrite (CuFeS2), and sphalerite (ZnS), with additions of galena (PbS) and alabandite (MnS). ZnS and MnS have a unique ability to store radiation energy, e.g. from ultraviolet light. When replicating molecules were originating, the primordial atmospheric pressure was high enough (>100 bar) to precipitate near the Earth’s surface, and ultraviolet irradiation was 10 to 100 times more intense than now; hence the photosynthetic properties mediated by ZnS provided the right energy conditions for the synthesis of informational and metabolic molecules and the selection of photostable nucleobases.[224][226]
The Zn-world theory has been filled out with evidence for the ionic constitution of the interior of the first protocells. In 1926, the Canadian biochemist Archibald Macallum noted the resemblance of body fluids such as blood and lymph to seawater;[227] however, the inorganic composition of all cells differ from that of modern seawater, which led Mulkidjanian and colleagues to reconstruct the «hatcheries» of the first cells combining geochemical analysis with phylogenomic scrutiny of the inorganic ion requirements of modern cells. The authors conclude that ubiquitous, and by inference primordial, proteins and functional systems show affinity to and functional requirement for K+, Zn2+, Mn2+, and [PO
4]3−
. Geochemical reconstruction shows that this ionic composition could not have existed in the ocean but is compatible with inland geothermal systems. In the oxygen-depleted, CO2-dominated primordial atmosphere, the chemistry of water condensates near geothermal fields would resemble the internal milieu of modern cells. Therefore, precellular evolution may have taken place in shallow «Darwin ponds» lined with porous silicate minerals mixed with metal sulfides and enriched in K+, Zn2+, and phosphorus compounds.[228][229]
Homochirality[edit]
Many biomolecules, such as L-glutamic acid, are asymmetric, and occur in living systems in only one of the two possible forms, in the case of amino acids the left-handed form. Prebiotic chemistry would produce both forms, creating a puzzle for abiogenesis researchers.[230]
Homochirality is the geometric uniformity of materials composed of chiral (non-mirror-symmetric) units. Living organisms use molecules that have the same chirality (handedness): with almost no exceptions,[231] amino acids are left-handed while nucleotides and sugars are right-handed. Chiral molecules can be synthesized, but in the absence of a chiral source or a chiral catalyst, they are formed in a 50/50 (racemic) mixture of both forms. Known mechanisms for the production of non-racemic mixtures from racemic starting materials include: asymmetric physical laws, such as the electroweak interaction; asymmetric environments, such as those caused by circularly polarized light, quartz crystals, or the Earth’s rotation, statistical fluctuations during racemic synthesis,[230] and spontaneous symmetry breaking.[232][233][234] Once established, chirality would be selected for.[235] A small bias (enantiomeric excess) in the population can be amplified into a large one by asymmetric autocatalysis, such as in the Soai reaction.[236] In asymmetric autocatalysis, the catalyst is a chiral molecule, which means that a chiral molecule is catalyzing its own production. An initial enantiomeric excess, such as can be produced by polarized light, then allows the more abundant enantiomer to outcompete the other.[237] Homochirality may have started in outer space, as on the Murchison meteorite the amino acid L-alanine is more than twice as frequent as its D form, and L-glutamic acid is more than three times as abundant as its D counterpart.[238][239] Amino acids from meteorites show a left-handed bias, whereas sugars show a predominantly right-handed bias, as found in living organisms, suggesting an abiogenic origin of these compounds.[240]
See also[edit]
- Formamide-based prebiotic chemistry – Scientific efforts aimed at reconstructing the beginnings of life
- GADV-protein world hypothesis – A hypothetical stage of abiogenesis
- Genetic recombination – Production of offspring with combinations of traits that differ from those found in either parent
- Shadow biosphere – Hypothetical biosphere of Earth
References[edit]
Explanatory footnotes[edit]
- ^ The reactions are:
- FeS + H2S → FeS2 + 2H+ + 2e−
- FeS + H2S + CO2 → FeS2 + HCOOH
- ^ The reactions are:
Reaction 1: Fayalite + water → magnetite + aqueous silica + hydrogen- 3Fe2SiO4 + 2H2O → 2Fe3O4 + 3SiO2 + 2H2
Reaction 2: Forsterite + aqueous silica → serpentine
- 3Mg2SiO4 + SiO2 + 4H2O → 2Mg3Si2O5(OH)4
Reaction 3: Forsterite + water → serpentine + brucite
- 2Mg2SiO4 + 3H2O → Mg3Si2O5(OH)4 + Mg(OH)2
Reaction 3 describes the hydration of olivine with water only to yield serpentine and Mg(OH)2 (brucite). Serpentine is stable at high pH in the presence of brucite like calcium silicate hydrate, (C-S-H) phases formed along with portlandite (Ca(OH)2) in hardened Portland cement paste after the hydration of belite (Ca2SiO4), the artificial calcium equivalent of forsterite.
Analogy of reaction 3 with belite hydration in ordinary Portland cement: Belite + water → C-S-H phase + portlandite
- 2 Ca2SiO4 + 4 H2O → 3 CaO · 2 SiO2 · 3 H2O + Ca(OH)2
Citations[edit]
- ^ a b Walker, Sara I.; Packard, N.; Cody, G. D. (13 November 2017). «Re-conceptualizing the origins of life». Philosophical Transactions of the Royal Society A. 375 (2109): 20160337. Bibcode:2017RSPTA.37560337W. doi:10.1098/rsta.2016.0337. PMC 5686397. PMID 29133439.
- ^ a b c «NASA Astrobiology Strategy» (PDF). NASA. 2015. Archived from the original (PDF) on 22 December 2016. Retrieved 24 September 2017.
- ^ Trifonov, Edward N. (17 March 2011). «Vocabulary of Definitions of Life Suggests a Definition». Journal of Biomolecular Structure and Dynamics. 29 (2): 259–266. doi:10.1080/073911011010524992. PMID 21875147. S2CID 38476092.
- ^ Voytek, Mary A. (6 March 2021). «About Life Detection». NASA. Retrieved 8 March 2021.
- ^ a b Witzany, Guenther (2016). «Crucial steps to life: From chemical reactions to code using agents» (PDF). BioSystems. 140: 49–57. doi:10.1016/j.biosystems.2015.12.007. PMID 26723230. S2CID 30962295.
- ^ a b Howell, Elizabeth (8 December 2014). «How Did Life Become Complex, And Could It Happen Beyond Earth?». Astrobiology Magazine. Archived from the original on 15 February 2018. Retrieved 14 April 2022.
{{cite web}}: CS1 maint: unfit URL (link) - ^ Oparin, Aleksandr Ivanovich (2003) [1938]. The Origin of Life. Translated by Morgulis, Sergius (2 ed.). Mineola, New York: Courier. ISBN 978-0486495224.
- ^ a b Peretó, Juli (2005). «Controversies on the origin of life» (PDF). International Microbiology. 8 (1): 23–31. PMID 15906258. Archived from the original (PDF) on 24 August 2015. Retrieved 1 June 2015.
- ^ Compare: Scharf, Caleb; et al. (18 December 2015). «A Strategy for Origins of Life Research». Astrobiology. 15 (12): 1031–1042. Bibcode:2015AsBio..15.1031S. doi:10.1089/ast.2015.1113. PMC 4683543. PMID 26684503.
What do we mean by the origins of life (OoL)? … Since the early 20th century the phrase OoL has been used to refer to the events that occurred during the transition from non-living to living systems on Earth, i.e., the origin of terrestrial biology (Oparin, 1924; Haldane, 1929). The term has largely replaced earlier concepts such as abiogenesis (Kamminga, 1980; Fry, 2000).
- ^ a b c Weiss, M. C.; Sousa, F. L.; Mrnjavac, N.; Neukirchen, S.; Roettger, M.; Nelson-Sathi, S.; Martin, W.F. (2016). «The physiology and habitat of the last universal common ancestor» (PDF). Nature Microbiology. 1 (9): 16116. doi:10.1038/NMICROBIOL.2016.116. PMID 27562259. S2CID 2997255.
- ^ Tirard, Stephane (20 April 2015). Abiogenesis – Definition. Encyclopedia of Astrobiology. p. 1. doi:10.1007/978-3-642-27833-4_2-4. ISBN 978-3-642-27833-4.
Thomas Huxley (1825–1895) used the term abiogenesis in an important text published in 1870. He strictly made the difference between spontaneous generation, which he did not accept, and the possibility of the evolution of matter from inert to living, without any influence of life. … Since the end of the nineteenth century, evolutive abiogenesis means increasing complexity and evolution of matter from inert to living state in the abiotic context of evolution of primitive Earth.
- ^ Graham, Robert W. (February 1990). «Extraterrestrial Life in the Universe» (PDF). NASA (NASA Technical Memorandum 102363). Lewis Research Center, Cleveland, Ohio. Archived (PDF) from the original on 3 September 2014. Retrieved 2 June 2015.
- ^ Altermann 2009, p. xvii
- ^ Oparin 1953, p. vi
- ^ Warmflash, David; Warmflash, Benjamin (November 2005). «Did Life Come from Another World?». Scientific American. 293 (5): 64–71. Bibcode:2005SciAm.293e..64W. doi:10.1038/scientificamerican1105-64. PMID 16318028.
- ^ Yarus 2010, p. 47
- ^ Ward, Peter; Kirschvink, Joe (2015). A New History of Life: the radical discoveries about the origins and evolution of life on earth. Bloomsbury Press. pp. 39–40. ISBN 978-1608199105.
- ^ Sheldon 2005
- ^ Lennox 2001, pp. 229–258
- ^ a b Bernal 1967
- ^ Balme, D. M. (1962). «Development of Biology in Aristotle and Theophrastus: Theory of Spontaneous Generation». Phronesis. 7 (1–2): 91–104. doi:10.1163/156852862X00052.
- ^ Ross 1652
- ^ Dobell 1960
- ^ Bondeson 1999
- ^ Levine, R.; Evers, C. «The Slow Death of Spontaneous Generation (1668-1859)». Archived from the original on 26 April 2008. Retrieved 18 April 2013.
- ^ Oparin 1953, p. 196
- ^ Tyndall 1905, IV, XII (1876), XIII (1878)
- ^ Horneck, Gerda; Klaus, David M.; Mancinelli, Rocco L. (March 2010). «Space Microbiology». Microbiology and Molecular Biology Reviews. 74 (1): 121–156. Bibcode:2010MMBR…74..121H. doi:10.1128/MMBR.00016-09. PMC 2832349. PMID 20197502.
- ^ Wickramasinghe, Chandra (2011). «Bacterial morphologies supporting cometary panspermia: a reappraisal». International Journal of Astrobiology. 10 (1): 25–30. Bibcode:2011IJAsB..10…25W. CiteSeerX 10.1.1.368.4449. doi:10.1017/S1473550410000157. S2CID 7262449.
- ^ Rampelotto, P. H. (2010). «Panspermia: A promising field of research». In: Astrobiology Science Conference. Abs 5224.
- ^ Chang, Kenneth (12 September 2016). «Visions of Life on Mars in Earth’s Depths». The New York Times. Archived from the original on 12 September 2016. Retrieved 12 September 2016.
- ^ «Letter no. 7471, Charles Darwin to Joseph Dalton Hooker, 1 February (1871)». Darwin Correspondence Project. Retrieved 7 July 2020.
- ^ Priscu, John C. «Origin and Evolution of Life on a Frozen Earth». Arlington County, Virginia: National Science Foundation. Archived from the original on 18 December 2013. Retrieved 1 March 2014.
- ^ Marshall, Michael (11 November 2020). «Charles Darwin’s hunch about early life was probably right». BBC News. Retrieved 11 November 2020.
- ^ Bahadur, Krishna (1973). «Photochemical Formation of Self–sustaining Coacervates» (PDF). Proceedings of the Indian National Science Academy. 39 (4): 455–467. doi:10.1016/S0044-4057(75)80076-1. PMID 1242552. Archived from the original (PDF) on 19 October 2013.
- ^ Bahadur, Krishna (1975). «Photochemical Formation of Self-Sustaining Coacervates». Zentralblatt für Bakteriologie, Parasitenkunde, Infektionskrankheiten und Hygiene (Central Journal for Bacteriology, Parasitology, Infectious Diseases and Hygiene). 130 (3): 211–218. doi:10.1016/S0044-4057(75)80076-1. OCLC 641018092. PMID 1242552.
- ^ Bryson 2004, pp. 300–302
- ^ Bernal 1951
- ^ Martin, William F. (January 2003). «On the origins of cells: a hypothesis for the evolutionary transitions from abiotic geochemistry to chemoautotrophic prokaryotes, and from prokaryotes to nucleated cells». Phil. Trans. R. Soc. Lond. A. 358 (1429): 59–83. doi:10.1098/rstb.2002.1183. PMC 1693102. PMID 12594918.
- ^ Bernal, John Desmond (September 1949). «The Physical Basis of Life». Proceedings of the Physical Society, Section A. 62 (9): 537–558. Bibcode:1949PPSA…62..537B. doi:10.1088/0370-1298/62/9/301. S2CID 83754271.
- ^ Miller, Stanley L. (15 May 1953). «A Production of Amino Acids Under Possible Primitive Earth Conditions». Science. 117 (3046): 528–529. Bibcode:1953Sci…117..528M. doi:10.1126/science.117.3046.528. PMID 13056598.
- ^ Parker, Eric T.; Cleaves, Henderson J.; Dworkin, Jason P.; et al. (5 April 2011). «Primordial synthesis of amines and amino acids in a 1958 Miller H2S-rich spark discharge experiment». PNAS. 108 (14): 5526–5531. Bibcode:2011PNAS..108.5526P. doi:10.1073/pnas.1019191108. PMC 3078417. PMID 21422282.
- ^ Bernal 1967, p. 143
- ^ a b Cleaves, H. James; Chalmers, John H.; Lazcano, Antonio; et al. (April 2008). «A Reassessment of Prebiotic Organic Synthesis in Neutral Planetary Atmospheres». Origins of Life and Evolution of Biospheres. 38 (2): 105–115. Bibcode:2008OLEB…38..105C. doi:10.1007/s11084-007-9120-3. PMID 18204914. S2CID 7731172.
- ^ Chyba, Christopher F. (13 May 2005). «Rethinking Earth’s Early Atmosphere». Science. 308 (5724): 962–963. doi:10.1126/science.1113157. PMID 15890865. S2CID 93303848.
- ^ Barton et al. 2007, pp. 93–95
- ^ Bada & Lazcano 2009, pp. 56–57
- ^ Bada, Jeffrey L.; Lazcano, Antonio (2 May 2003). «Prebiotic Soup – Revisiting the Miller Experiment» (PDF). Science. 300 (5620): 745–746. doi:10.1126/science.1085145. PMID 12730584. S2CID 93020326. Archived (PDF) from the original on 4 March 2016. Retrieved 13 June 2015.
- ^ Marigo, Paola; et al. (6 July 2020). «Carbon star formation as seen through the non-monotonic initial–final mass relation». Nature Astronomy. 152 (11): 1102–1110. arXiv:2007.04163. Bibcode:2020NatAs…4.1102M. doi:10.1038/s41550-020-1132-1. S2CID 220403402.
- ^ «WMAP- Life in the Universe».
- ^ «Formation of Solar Systems: Solar Nebular Theory». University of Massachusetts Amherst. Retrieved 27 September 2019.
- ^ «Age of the Earth». United States Geological Survey. 9 July 2007. Archived from the original on 23 December 2005. Retrieved 10 January 2006.
- ^ Dalrymple 2001, pp. 205–221
- ^ Fesenkov 1959, p. 9
- ^ Kasting, James F. (12 February 1993). «Earth’s Early Atmosphere» (PDF). Science. 259 (5097): 920–926. Bibcode:1993Sci…259..920K. doi:10.1126/science.11536547. PMID 11536547. S2CID 21134564. Archived from the original (PDF) on 10 October 2015. Retrieved 28 July 2015.
- ^ Morse, John (September 1998). «Hadean Ocean Carbonate Geochemistry». Aquatic Geochemistry. 4 (3/4): 301–319. Bibcode:1998MinM…62.1027M. doi:10.1023/A:1009632230875. S2CID 129616933.
- ^ a b c d e f g h Follmann, Hartmut; Brownson, Carol (November 2009). «Darwin’s warm little pond revisited: from molecules to the origin of life». Naturwissenschaften. 96 (11): 1265–1292. Bibcode:2009NW…..96.1265F. doi:10.1007/s00114-009-0602-1. PMID 19760276. S2CID 23259886.
- ^ Morse, John W.; MacKenzie, Fred T. (1998). «Hadean Ocean Carbonate Geochemistry». Aquatic Geochemistry. 4 (3–4): 301–319. Bibcode:1998MinM…62.1027M. doi:10.1023/A:1009632230875. S2CID 129616933.
- ^ Wilde, Simon A.; Valley, John W.; Peck, William H.; Graham, Colin M. (11 January 2001). «Evidence from detrital zircons for the existence of continental crust and oceans on the Earth 4.4 Gyr ago» (PDF). Nature. 409 (6817): 175–178. Bibcode:2001Natur.409..175W. doi:10.1038/35051550. PMID 11196637. S2CID 4319774. Archived (PDF) from the original on 5 June 2015. Retrieved 3 June 2015.
- ^ Rosing, Minik T.; Bird, Dennis K.; Sleep, Norman H.; et al. (22 March 2006). «The rise of continents – An essay on the geologic consequences of photosynthesis». Palaeogeography, Palaeoclimatology, Palaeoecology. 232 (2–4): 99–113. Bibcode:2006PPP…232…99R. doi:10.1016/j.palaeo.2006.01.007. Archived (PDF) from the original on 14 July 2015. Retrieved 8 June 2015.
- ^ a b c Dodd, Matthew S.; Papineau, Dominic; Grenne, Tor; et al. (1 March 2017). «Evidence for early life in Earth’s oldest hydrothermal vent precipitates». Nature. 543 (7643): 60–64. Bibcode:2017Natur.543…60D. doi:10.1038/nature21377. PMID 28252057. Archived from the original on 8 September 2017. Retrieved 2 March 2017.
- ^ Gomes, Rodney; Levison, Hal F.; Tsiganis, Kleomenis; Morbidelli, Alessandro (26 May 2005). «Origin of the cataclysmic Late Heavy Bombardment period of the terrestrial planets». Nature. 435 (7041): 466–469. Bibcode:2005Natur.435..466G. doi:10.1038/nature03676. PMID 15917802.
- ^ Sleep, Norman H.; Zahnle, Kevin J.; Kasting, James F.; et al. (9 November 1989). «Annihilation of ecosystems by large asteroid impacts on early Earth». Nature. 342 (6246): 139–142. Bibcode:1989Natur.342..139S. doi:10.1038/342139a0. PMID 11536616. S2CID 1137852.
- ^ Chyba, Christopher; Sagan, Carl (9 January 1992). «Endogenous production, exogenous delivery and impact-shock synthesis of organic molecules: an inventory for the origins of life». Nature. 355 (6356): 125–132. Bibcode:1992Natur.355..125C. doi:10.1038/355125a0. PMID 11538392. S2CID 4346044.
- ^ Furukawa, Yoshihiro; Sekine, Toshimori; Oba, Masahiro; et al. (January 2009). «Biomolecule formation by oceanic impacts on early Earth». Nature Geoscience. 2 (1): 62–66. Bibcode:2009NatGe…2…62F. doi:10.1038/NGEO383.
- ^ Maher, Kevin A.; Stevenson, David J. (18 February 1988). «Impact frustration of the origin of life». Nature. 331 (6157): 612–614. Bibcode:1988Natur.331..612M. doi:10.1038/331612a0. PMID 11536595. S2CID 4284492.
- ^ Mann, Adam (24 January 2018). «Bashing holes in the tale of Earth’s troubled youth». Nature. 553 (7689): 393–395. Bibcode:2018Natur.553..393M. doi:10.1038/d41586-018-01074-6.
- ^ Davies 1999, p. 155
- ^ Bock & Goode 1996
- ^ Schopf, J. William; Kudryavtsev, Anatoliy B.; Czaja, Andrew D.; Tripathi, Abhishek B. (5 October 2007). «Evidence of Archean life: Stromatolites and microfossils». Precambrian Research. 158 (3–4): 141–155. Bibcode:2007PreR..158..141S. doi:10.1016/j.precamres.2007.04.009.
- ^ Schopf, J. William (29 June 2006). «Fossil evidence of Archaean life». Philosophical Transactions of the Royal Society B. 361 (1470): 869–885. doi:10.1098/rstb.2006.1834. PMC 1578735. PMID 16754604.
- ^ Raven & Johnson 2002, p. 68
- ^ Djokic, Tara; Van Kranendonk, Martin J.; Campbell, Kathleen A.; Walter, Malcolm R.; Ward, Colin R. (9 May 2017). «Earliest signs of life on land preserved in ca. 3.5 Gao hot spring deposits». Nature Communications. 8: 15263. Bibcode:2017NatCo…815263D. doi:10.1038/ncomms15263. PMC 5436104. PMID 28486437.
- ^ Schopf, J. William; Kitajima, Kouki; Spicuzza, Michael J.; Kudryavtsev, Anatolly B.; Valley, John W. (2017). «SIMS analyses of the oldest known assemblage of microfossils document their taxon-correlated carbon isotope compositions». PNAS. 115 (1): 53–58. Bibcode:2018PNAS..115…53S. doi:10.1073/pnas.1718063115. PMC 5776830. PMID 29255053.
- ^ Tyrell, Kelly April (18 December 2017). «Oldest fossils ever found show life on Earth began before 3.5 billion years ago». University of Wisconsin-Madison. Retrieved 18 December 2017.
- ^ Ohtomo, Yoko; Kakegawa, Takeshi; Ishida, Akizumi; et al. (January 2014). «Evidence for biogenic graphite in early Archaean Isua metasedimentary rocks». Nature Geoscience. 7 (1): 25–28. Bibcode:2014NatGe…7…25O. doi:10.1038/ngeo2025.
- ^ Noffke, Nora; Christian, Daniel; Wacey, David; Hazen, Robert M. (16 November 2013). «Microbially Induced Sedimentary Structures Recording an Ancient Ecosystem in the ca. 3.48 Gyo Dresser Formation, Pilbara, Western Australia». Astrobiology. 13 (12): 1103–1124. Bibcode:2013AsBio..13.1103N. doi:10.1089/ast.2013.1030. PMC 3870916. PMID 24205812.
- ^ Davies 1999
- ^ Hassenkam, T.; Andersson, M. P.; Dalby, K. N.; Mackenzie, D.M.A.; Rosing, M.T. (2017). «Elements of Eoarchean life trapped in mineral inclusions». Nature. 548 (7665): 78–81. Bibcode:2017Natur.548…78H. doi:10.1038/nature23261. PMID 28738409. S2CID 205257931.
- ^ O’Donoghue, James (21 August 2011). «Oldest reliable fossils show early life was a beach». New Scientist. 211: 13. doi:10.1016/S0262-4079(11)62064-2. Archived from the original on 30 June 2015.
- ^ Wacey, David; Kilburn, Matt R.; Saunders, Martin; et al. (October 2011). «Microfossils of sulphur-metabolizing cells in 3.4-billion-year-old rocks of Western Australia». Nature Geoscience. 4 (10): 698–702. Bibcode:2011NatGe…4..698W. doi:10.1038/ngeo1238.
- ^ Bell, Elizabeth A.; Boehnike, Patrick; Harrison, T. Mark; et al. (19 October 2015). «Potentially biogenic carbon preserved in a 4.1 billion-year-old zircon». PNAS. 112 (47): 14518–14521. Bibcode:2015PNAS..11214518B. doi:10.1073/pnas.1517557112. PMC 4664351. PMID 26483481. Early edition, published online before print.
- ^ Baumgartner, Rafael; Van Kranendonk, Martin; Wacey, David; et al. (2019). «Nano−porous pyrite and organic matter in 3.5-billion-year-old stromatolites record primordial life» (PDF). Geology. 47 (11): 1039–1043. Bibcode:2019Geo….47.1039B. doi:10.1130/G46365.1. S2CID 204258554.
- ^ Djokic, Tara; Van Kranendonk, Martin; Cambell, Kathleen; Walter, Malcolm (2017). «Earliest signs of life on land preserved in ca. 3.5 Ga hot spring deposits». Nature Communications. 3.
- ^ Landau, Elizabeth (12 October 2016). «Building Blocks of Life’s Building Blocks Come From Starlight». NASA. Archived from the original on 13 October 2016. Retrieved 13 October 2016.
- ^ a b Ehrenfreund, Pascale; Cami, Jan (December 2010). «Cosmic carbon chemistry: from the interstellar medium to the early Earth». Cold Spring Harbor Perspectives in Biology. 2 (12): a002097. doi:10.1101/cshperspect.a002097. PMC 2982172. PMID 20554702.
- ^ Geballe, Thomas R.; Najarro, Francisco; Figer, Donald F.; et al. (10 November 2011). «Infrared diffuse interstellar bands in the Galactic Centre region». Nature. 479 (7372): 200–202. arXiv:1111.0613. Bibcode:2011Natur.479..200G. doi:10.1038/nature10527. PMID 22048316. S2CID 17223339.
- ^ Klyce 2001
- ^ a b c d Hoover, Rachel (21 February 2014). «Need to Track Organic Nano-Particles Across the Universe? NASA’s Got an App for That». Ames Research Center. NASA. Archived from the original on 6 September 2015. Retrieved 22 June 2015.
- ^ Goncharuk, Vladislav V.; Zui, O. V. (February 2015). «Water and carbon dioxide as the main precursors of organic matter on Earth and in space». Journal of Water Chemistry and Technology. 37 (1): 2–3. doi:10.3103/S1063455X15010026. S2CID 97965067.
- ^ Abou Mrad, Ninette; Vinogradoff, Vassilissa; Duvernay, Fabrice; et al. (2015). «Laboratory experimental simulations: Chemical evolution of the organic matter from interstellar and cometary ice analogs». Bulletin de la Société Royale des Sciences de Liège. 84: 21–32. Bibcode:2015BSRSL..84…21A. Archived from the original on 13 April 2015. Retrieved 6 April 2015.
- ^ Oba, Yasuhiro; et al. (26 April 2022). «Identifying the wide diversity of extraterrestrial purine and pyrimidine nucleobases in carbonaceous meteorites». Nature Communications. 13 (2008): 2008. Bibcode:2022NatCo..13.2008O. doi:10.1038/s41467-022-29612-x. PMC 9042847. PMID 35473908. S2CID 248402205.
- ^ «‘Life chemical’ detected in comet». BBC News. London. 18 August 2009. Archived from the original on 25 May 2015. Retrieved 23 June 2015.
- ^ Thompson, William Reid; Murray, B. G.; Khare, Bishun Narain; Sagan, Carl (30 December 1987). «Coloration and darkening of methane clathrate and other ices by charged particle irradiation: Applications to the outer solar system». Journal of Geophysical Research. 92 (A13): 14933–14947. Bibcode:1987JGR….9214933T. doi:10.1029/JA092iA13p14933. PMID 11542127.
- ^ Goldman, Nir; Tamblyn, Isaac (20 June 2013). «Prebiotic Chemistry within a Simple Impacting Icy Mixture». Journal of Physical Chemistry A. 117 (24): 5124–5131. Bibcode:2013JPCA..117.5124G. doi:10.1021/jp402976n. PMID 23639050. S2CID 5144843.
- ^ «NASA Ames PAH IR Spectroscopic Database». NASA. Archived from the original on 29 June 2015. Retrieved 17 June 2015.
- ^ a b c Hudgins, Douglas M.; Bauschlicher, Charles W. Jr.; Allamandola, Louis J. (10 October 2005). «Variations in the Peak Position of the 6.2 μm Interstellar Emission Feature: A Tracer of N in the Interstellar Polycyclic Aromatic Hydrocarbon Population». The Astrophysical Journal. 632 (1): 316–332. Bibcode:2005ApJ…632..316H. CiteSeerX 10.1.1.218.8786. doi:10.1086/432495. S2CID 7808613.
- ^ a b c Des Marais, David J.; Allamandola, Louis J.; Sandford, Scott; et al. (2009). «Cosmic Distribution of Chemical Complexity». Ames Research Center. Mountain View, California: NASA. Archived from the original on 27 February 2014. Retrieved 24 June 2015.
- ^ a b Carey, Bjorn (18 October 2005). «Life’s Building Blocks ‘Abundant in Space’«. Space.com. Watsonville, California: Imaginova. Archived from the original on 26 June 2015. Retrieved 23 June 2015.
- ^ García-Hernández, Domingo. A.; Manchado, Arturo; García-Lario, Pedro; et al. (20 November 2010). «Formation of Fullerenes in H-Containing Planetary Nebulae». The Astrophysical Journal Letters. 724 (1): L39–L43. arXiv:1009.4357. Bibcode:2010ApJ…724L..39G. doi:10.1088/2041-8205/724/1/L39. S2CID 119121764.
- ^ d’Ischia, Marco; Manini, Paola; Moracci, Marco; et al. (21 August 2019). «Astrochemistry and Astrobiology: Materials Science in Wonderland?». International Journal of Molecular Sciences. 20 (17): 4079. doi:10.3390/ijms20174079. PMC 6747172. PMID 31438518.
- ^ Gudipati, Murthy S.; Yang, Rui (1 September 2012). «In-situ Probing of Radiation-induced Processing of Organics in Astrophysical Ice Analogs – Novel Laser Desorption Laser Ionization Time-of-flight Mass Spectroscopic Studies». The Astrophysical Journal Letters. 756 (1): L24. Bibcode:2012ApJ…756L..24G. doi:10.1088/2041-8205/756/1/L24. S2CID 5541727.
- ^ a b Gallori, Enzo (June 2011). «Astrochemistry and the origin of genetic material». Rendiconti Lincei. 22 (2): 113–118. doi:10.1007/s12210-011-0118-4. S2CID 96659714. «Paper presented at the Symposium ‘Astrochemistry: molecules in space and time’ (Rome, 4–5 November 2010), sponsored by Fondazione ‘Guido Donegani’, Accademia Nazionale dei Lincei.»
- ^ Martins, Zita (February 2011). «Organic Chemistry of Carbonaceous Meteorites». Elements. 7 (1): 35–40. doi:10.2113/gselements.7.1.35.
- ^ Martins, Zita; Botta, Oliver; Fogel, Marilyn L.; et al. (15 June 2008). «Extraterrestrial nucleobases in the Murchison meteorite». Earth and Planetary Science Letters. 270 (1–2): 130–136. arXiv:0806.2286. Bibcode:2008E&PSL.270..130M. doi:10.1016/j.epsl.2008.03.026. S2CID 14309508.
- ^ Callahan, Michael P.; Smith, Karen E.; Cleaves, H. James, II; et al. (23 August 2011). «Carbonaceous meteorites contain a wide range of extraterrestrial nucleobases». PNAS. 108 (34): 13995–13998. Bibcode:2011PNAS..10813995C. doi:10.1073/pnas.1106493108. PMC 3161613. PMID 21836052.
- ^ Steigerwald, John (8 August 2011). «NASA Researchers: DNA Building Blocks Can Be Made in Space». Goddard Space Flight Center. NASA. Archived from the original on 23 June 2015. Retrieved 23 June 2015.
- ^ Kwok, Sun; Zhang, Yong (3 November 2011). «Mixed aromatic–aliphatic organic nanoparticles as carriers of unidentified infrared emission features». Nature. 479 (7371): 80–83. Bibcode:2011Natur.479…80K. doi:10.1038/nature10542. PMID 22031328. S2CID 4419859.
- ^ Jørgensen, Jes K.; Favre, Cécile; Bisschop, Suzanne E.; et al. (2012). «Detection of the simplest sugar, glycolaldehyde, in a solar-type protostar with ALMA» (PDF). The Astrophysical Journal Letters. 757 (1): L4. arXiv:1208.5498. Bibcode:2012ApJ…757L…4J. doi:10.1088/2041-8205/757/1/L4. S2CID 14205612. Archived (PDF) from the original on 24 September 2015. Retrieved 23 June 2015.
- ^ Furukawa, Yoshihiro; Chikaraishi, Yoshito; Ohkouchi, Naohiko; et al. (13 November 2019). «Extraterrestrial ribose and other sugars in primitive meteorites». PNAS. 116 (49): 24440–24445. Bibcode:2019PNAS..11624440F. doi:10.1073/pnas.1907169116. PMC 6900709. PMID 31740594.
- ^ Oró, Joan; Kimball, Aubrey P. (February 1962). «Synthesis of purines under possible primitive earth conditions: II. Purine intermediates from hydrogen cyanide». Archives of Biochemistry and Biophysics. 96 (2): 293–313. doi:10.1016/0003-9861(62)90412-5. PMID 14482339.
- ^ Cleaves II, Henderson (2010). «The origin of the biologically coded amino acids». Journal of Theoretical Biology. 263 (4): 490–498. Bibcode:2010JThBi.263..490C. doi:10.1016/j.jtbi.2009.12.014. PMID 20034500.
- ^ Breslow, R. (1959). «On the Mechanism of the Formose Reaction». Tetrahedron Letters. 1 (21): 22–26. doi:10.1016/S0040-4039(01)99487-0.
- ^ Oró, Joan (16 September 1961). «Mechanism of Synthesis of Adenine from Hydrogen Cyanide under Possible Primitive Earth Conditions». Nature. 191 (4794): 1193–1194. Bibcode:1961Natur.191.1193O. doi:10.1038/1911193a0. PMID 13731264. S2CID 4276712.
- ^ a b Saladino, Raffaele; Crestini, Claudia; Pino, Samanta; et al. (March 2012). «Formamide and the origin of life» (PDF). Physics of Life Reviews. 9 (1): 84–104. Bibcode:2012PhLRv…9…84S. doi:10.1016/j.plrev.2011.12.002. hdl:2108/85168. PMID 22196896.
- ^ a b Saladino, Raffaele; Botta, Giorgia; Pino, Samanta; et al. (July 2012). «From the one-carbon amide formamide to RNA all the steps are prebiotically possible». Biochimie. 94 (7): 1451–1456. doi:10.1016/j.biochi.2012.02.018. hdl:11573/515604. PMID 22738728.
- ^ Marlaire, Ruth, ed. (3 March 2015). «NASA Ames Reproduces the Building Blocks of Life in Laboratory». Ames Research Center. NASA. Archived from the original on 5 March 2015. Retrieved 5 March 2015.
- ^ Ferus, Martin; Nesvorný, David; Šponer, Jiří; et al. (2015). «High-energy chemistry of formamide: A unified mechanism of nucleobase formation». PNAS. 112 (3): 657–662. Bibcode:2015PNAS..112..657F. doi:10.1073/pnas.1412072111. PMC 4311869. PMID 25489115.
- ^ Basile, Brenda; Lazcano, Antonio; Oró, Joan (1984). «Prebiotic syntheses of purines and pyrimidines». Advances in Space Research. 4 (12): 125–131. Bibcode:1984AdSpR…4l.125B. doi:10.1016/0273-1177(84)90554-4. PMID 11537766.
- ^ Orgel, Leslie E. (August 2004). «Prebiotic Adenine Revisited: Eutectics and Photochemistry». Origins of Life and Evolution of Biospheres. 34 (4): 361–369. Bibcode:2004OLEB…34..361O. doi:10.1023/B:ORIG.0000029882.52156.c2. PMID 15279171. S2CID 4998122.
- ^ Robertson, Michael P.; Miller, Stanley L. (29 June 1995). «An efficient prebiotic synthesis of cytosine and uracil». Nature. 375 (6534): 772–774. Bibcode:1995Natur.375..772R. doi:10.1038/375772a0. PMID 7596408. S2CID 4351012.
- ^ Fox, Douglas (February 2008). «Did Life Evolve in Ice?». Discover. Archived from the original on 30 June 2008. Retrieved 3 July 2008.
- ^ Levy, Matthew; Miller, Stanley L.; Brinton, Karen; Bada, Jeffrey L. (June 2000). «Prebiotic Synthesis of Adenine and Amino Acids Under Europa-like Conditions». Icarus. 145 (2): 609–613. Bibcode:2000Icar..145..609L. doi:10.1006/icar.2000.6365. PMID 11543508.
- ^ Menor-Salván, César; Ruiz-Bermejo, Marta; Guzmán, Marcelo I.; et al. (20 April 2009). «Synthesis of Pyrimidines and Triazines in Ice: Implications for the Prebiotic Chemistry of Nucleobases». Chemistry: A European Journal. 15 (17): 4411–4418. doi:10.1002/chem.200802656. PMID 19288488.
- ^ Roy, Debjani; Najafian, Katayoun; von Ragué Schleyer, Paul (30 October 2007). «Chemical evolution: The mechanism of the formation of adenine under prebiotic conditions». PNAS. 104 (44): 17272–17277. Bibcode:2007PNAS..10417272R. doi:10.1073/pnas.0708434104. PMC 2077245. PMID 17951429.
- ^ Lancet, Doron (30 December 2014). «Systems Prebiology-Studies of the origin of Life». The Lancet Lab. Rehovot, Israel: Department of Molecular Genetics; Weizmann Institute of Science. Archived from the original on 26 June 2015. Retrieved 26 June 2015.
- ^ Segré, Daniel; Ben-Eli, Dafna; Deamer, David W.; Lancet, Doron (February 2001). «The Lipid World» (PDF). Origins of Life and Evolution of Biospheres. 31 (1–2): 119–145. Bibcode:2001OLEB…31..119S. doi:10.1023/A:1006746807104. PMID 11296516. S2CID 10959497. Archived (PDF) from the original on 26 June 2015.
- ^ a b c Chen, Irene A.; Walde, Peter (July 2010). «From Self-Assembled Vesicles to Protocells». Cold Spring Harbor Perspectives in Biology. 2 (7): a002170. doi:10.1101/cshperspect.a002170. PMC 2890201. PMID 20519344.
- ^ Eigen, Manfred; Schuster, Peter (November 1977). «The Hypercycle. A Principle of Natural Self-Organization. Part A: Emergence of the Hypercycle» (PDF). Naturwissenschaften. 64 (11): 541–65. Bibcode:1977NW…..64..541E. doi:10.1007/bf00450633. PMID 593400. S2CID 42131267. Archived from the original (PDF) on 3 March 2016.
- Eigen, Manfred; Schuster, Peter (1978). «The Hypercycle. A Principle of Natural Self-Organization. Part B: The Abstract Hypercycle» (PDF). Naturwissenschaften. 65 (1): 7–41. Bibcode:1978NW…..65….7E. doi:10.1007/bf00420631. S2CID 1812273. Archived from the original (PDF) on 3 March 2016.
- Eigen, Manfred; Schuster, Peter (July 1978). «The Hypercycle. A Principle of Natural Self-Organization. Part C: The Realistic Hypercycle» (PDF). Naturwissenschaften. 65 (7): 341–369. Bibcode:1978NW…..65..341E. doi:10.1007/bf00439699. S2CID 13825356. Archived from the original (PDF) on 16 June 2016.
- ^ Markovitch, Omer; Lancet, Doron (Summer 2012). «Excess Mutual Catalysis Is Required for Effective Evolvability». Artificial Life. 18 (3): 243–266. doi:10.1162/artl_a_00064. PMID 22662913. S2CID 5236043.
- ^ Tessera, Marc (2011). «Origin of Evolution versus Origin of Life: A Shift of Paradigm». International Journal of Molecular Sciences. 12 (6): 3445–3458. doi:10.3390/ijms12063445. PMC 3131571. PMID 21747687. Special Issue: «Origin of Life 2011»
- ^ Onsager, Lars (1931). «Reciprocal Relations in Irreversible Processes I and II». Physical Review. 37 (4): 405. Bibcode:1931PhRv…37..405O. doi:10.1103/PhysRev.37.405.
- ^ Onsager, Lars (1931). «Reciprocal Relations in Irreversible Processes I and II». Physical Review (38): 2265. doi:10.1103/PhysRev.38.2265.
- ^ Prigogine, Ilya (1967). An Introduction to the Thermodynamics of Irreversible Processes. New York: Wiley.
- ^ «Exploring Life’s Origins: Protocells». Exploring Life’s Origins: A Virtual Exhibit. Arlington County, Virginia: National Science Foundation. Archived from the original on 28 February 2014. Retrieved 18 March 2014.
- ^ a b c Chen, Irene A. (8 December 2006). «The Emergence of Cells During the Origin of Life». Science. 314 (5805): 1558–1559. doi:10.1126/science.1137541. PMID 17158315.
- ^ Zimmer, Carl (26 June 2004). «What Came Before DNA?». Discover. Archived from the original on 19 March 2014.
- ^ Shapiro, Robert (June 2007). «A Simpler Origin for Life». Scientific American. 296 (6): 46–53. Bibcode:2007SciAm.296f..46S. doi:10.1038/scientificamerican0607-46. PMID 17663224. Archived from the original on 14 June 2015.
- ^ Chang 2007
- ^ a b c d Lane, Nick (2015). The Vital Question: Why Is Life The Way It Is?. Profile Books. pp. 129–140. ISBN 978-1781250365.
- ^ Sharov, Alexei A.; Gordon, Richard (2018). «Life Before Earth». Habitability of the Universe Before Earth: Life Before Earth. Astrobiology Exploring Life on Earth and Beyond. Academic Press. pp. 265–296. doi:10.1016/B978-0-12-811940-2.00011-3. ISBN 9780128119402. S2CID 117048600.
- ^ Ladyman, J.; Lambert, J.; Weisner, K. B. (2013). «What is a Complex System?». European Journal of the Philosophy of Science. 3: 33–67. doi:10.1007/s13194-012-0056-8. S2CID 18787276.
- ^ Esposito, M.; Lindenberg, Katja; Van den Broeck, C. (2010). «Entropy production as correlation between system and reservoir». New Journal of Physics. 12 (1): 013013. arXiv:0908.1125. Bibcode:2010NJPh…12a3013E. doi:10.1088/1367-2630/12/1/013013. S2CID 26657293.
- ^ Bar-Nun, A.; Bar-Nun, N.; Bauer, S. H.; Sagan, Carl (24 April 1970). «Shock Synthesis of Amino Acids in Simulated Primitive Environments». Science. 168 (3930): 470–473. Bibcode:1970Sci…168..470B. doi:10.1126/science.168.3930.470. PMID 5436082. S2CID 42467812.
- ^ Anbar, Michael (27 September 1968). «Cavitation during Impact of Liquid Water on Water: Geochemical Implications». Science. 161 (3848): 1343–1344. Bibcode:1968Sci…161.1343A. doi:10.1126/science.161.3848.1343. PMID 17831346.
- ^ Dharmarathne, Leena; Grieser, Franz (7 January 2016). «Formation of Amino Acids on the Sonolysis of Aqueous Solutions Containing Acetic Acid, Methane, or Carbon Dioxide, in the Presence of Nitrogen Gas». The Journal of Physical Chemistry A. 120 (2): 191–199. Bibcode:2016JPCA..120..191D. doi:10.1021/acs.jpca.5b11858. PMID 26695890.
- ^ Patehebieke, Yeersen; Zhao, Ze-Run; Wang, Su; Xu, Hao-Xing; Chen, Qian-Qian; Wang, Xiao (2021). «Cavitation as a plausible driving force for the prebiotic formation of N9 purine nucleosides». Cell Reports Physical Science. 2 (3): 100375. Bibcode:2021CRPS….200375P. doi:10.1016/j.xcrp.2021.100375. S2CID 233662126.
- ^ Kalson, Natan-Haim; Furman, David; Zeiri, Yehuda (11 September 2017). «Cavitation-Induced Synthesis of Biogenic Molecules on Primordial Earth». ACS Central Science. 3 (9): 1041–1049. doi:10.1021/acscentsci.7b00325. PMC 5620973. PMID 28979946. S2CID 21409351.
- ^ Muller, Anthonie W. J. (1995). «Were the first organisms heat engines? A new model for biogenesis and the early evolution of biological energy conversion». Progress in Biophysics and Molecular Biology. 63 (2): 193–231. doi:10.1016/0079-6107(95)00004-7. PMID 7542789.
- ^ Muller, Anthonie W. J.; Schulze-Makuch, Dirk (2006). «Thermal energy and the origin of life». Origins of Life and Evolution of Biospheres. 36 (2): 77–189. Bibcode:2006OLEB…36..177M. doi:10.1007/s11084-005-9003-4. PMID 16642267. S2CID 22179552.
- ^ Junge, Wolfgang; Nelson, Nathan (2 June 2015). «ATP Synthase». Annual Review of Biochemistry. 84 (1): 631–657. doi:10.1146/annurev-biochem-060614-034124. PMID 25839341.
- ^ a b c Damer, Bruce; Deamer, David (1 April 2020). «The Hot Spring Hypothesis for an Origin of Life». Astrobiology. 20 (4): 429–452. Bibcode:2020AsBio..20..429D. doi:10.1089/ast.2019.2045. ISSN 1531-1074. PMC 7133448. PMID 31841362.
- ^ Benner, S. A.; Bell, E. A.; Biondi, E.; Brasser, R.; Carell, T.; Kim, H.-J.; Mojzsis, S. J.; Omran, A.; Pasek, M. A.; Trail, D. (2020). «When Did Life Likely Emerge on Earth in an RNA-First Process?». ChemSystemsChem. 2 (2). doi:10.1002/syst.201900035.
- ^ * Copley, Shelley D.; Smith, Eric; Morowitz, Harold J. (December 2007). «The origin of the RNA world: Co-evolution of genes and metabolism» (PDF). Bioorganic Chemistry. 35 (6): 430–443. doi:10.1016/j.bioorg.2007.08.001. PMID 17897696. Archived (PDF) from the original on 5 September 2013. Retrieved 8 June 2015.
The proposal that life on Earth arose from an RNA world is widely accepted.
- Orgel, Leslie E. (April 2003). «Some consequences of the RNA world hypothesis». Origins of Life and Evolution of Biospheres. 33 (2): 211–218. Bibcode:2003OLEB…33..211O. doi:10.1023/A:1024616317965. PMID 12967268. S2CID 32779859.
It now seems very likely that our familiar DNA/RNA/protein world was preceded by an RNA world…
- Robertson & Joyce 2012: «There is now strong evidence indicating that an RNA World did indeed exist before DNA- and protein-based life.»
- Neveu, Kim & Benner 2013: «[The RNA world’s existence] has broad support within the community today.»
- Orgel, Leslie E. (April 2003). «Some consequences of the RNA world hypothesis». Origins of Life and Evolution of Biospheres. 33 (2): 211–218. Bibcode:2003OLEB…33..211O. doi:10.1023/A:1024616317965. PMID 12967268. S2CID 32779859.
- ^ a b c Robertson, Michael P.; Joyce, Gerald F. (May 2012). «The origins of the RNA world». Cold Spring Harbor Perspectives in Biology. 4 (5): a003608. doi:10.1101/cshperspect.a003608. PMC 3331698. PMID 20739415.
- ^ a b c Cech, Thomas R. (July 2012). «The RNA Worlds in Context». Cold Spring Harbor Perspectives in Biology. 4 (7): a006742. doi:10.1101/cshperspect.a006742. PMC 3385955. PMID 21441585.
- ^ Pearce, Ben K. D.; Pudritz, Ralph E.; Semenov, Dmitry A.; Henning, Thomas K. (24 October 2017). «Origin of the RNA world: The fate of nucleobases in warm little ponds». PNAS. 114 (43): 11327–11332. arXiv:1710.00434. Bibcode:2017PNAS..11411327P. doi:10.1073/pnas.1710339114. PMC 5664528. PMID 28973920.
- ^ Yarus, Michael (April 2011). «Getting Past the RNA World: The Initial Darwinian Ancestor». Cold Spring Harbor Perspectives in Biology. 3 (4): a003590. doi:10.1101/cshperspect.a003590. PMC 3062219. PMID 20719875.
- ^ Voet & Voet 2004, p. 29
- ^ Fox, George.E. (9 June 2010). «Origin and evolution of the ribosome». Cold Spring Harbor Perspectives in Biology. 2 (9(a003483)): a003483. doi:10.1101/cshperspect.a003483. PMC 2926754. PMID 20534711.
- ^ Neveu, Marc; Kim, Hyo-Joong; Benner, Steven A. (22 April 2013). «The ‘Strong’ RNA World Hypothesis: Fifty Years Old». Astrobiology. 13 (4): 391–403. Bibcode:2013AsBio..13..391N. doi:10.1089/ast.2012.0868. PMID 23551238.
- ^ Gilbert, Walter (20 February 1986). «Origin of life: The RNA world». Nature. 319 (6055): 618. Bibcode:1986Natur.319..618G. doi:10.1038/319618a0. S2CID 8026658.
- ^ Orgel, Leslie E. (October 1994). «The origin of life on Earth». Scientific American. 271 (4): 76–83. Bibcode:1994SciAm.271d..76O. doi:10.1038/scientificamerican1094-76. PMID 7524147.
- ^ Lincoln, Tracey A.; Joyce, Gerald F. (27 February 2009). «Self-Sustained Replication of an RNA Enzyme». Science. 323 (5918): 1229–1232. Bibcode:2009Sci…323.1229L. doi:10.1126/science.1167856. PMC 2652413. PMID 19131595.
- ^ Joyce, Gerald F. (2009). «Evolution in an RNA world». Cold Spring Harbor Perspectives in Biology. 74 (Evolution: The Molecular Landscape): 17–23. doi:10.1101/sqb.2009.74.004. PMC 2891321. PMID 19667013.
- ^ Szostak, Jack W. (5 February 2015). «The Origins of Function in Biological Nucleic Acids, Proteins, and Membranes». Chevy Chase, Maryland: Howard Hughes Medical Institute. Archived from the original on 14 July 2015. Retrieved 16 June 2015.
- ^ Bernstein, Harris; Byerly, Henry C.; Hopf, Frederick A.; et al. (June 1983). «The Darwinian Dynamic». The Quarterly Review of Biology. 58 (2): 185–207. doi:10.1086/413216. JSTOR 2828805. S2CID 83956410.
- ^ Michod 1999
- ^ Palasek, Stan (23 May 2013). «Primordial RNA Replication and Applications in PCR Technology». arXiv:1305.5581v1 [q-bio.BM].
- ^ Vlassov, Alexander V.; Kazakov, Sergei A.; Johnston, Brian H.; et al. (August 2005). «The RNA World on Ice: A New Scenario for the Emergence of RNA Information». Journal of Molecular Evolution. 61 (2): 264–273. Bibcode:2005JMolE..61..264V. doi:10.1007/s00239-004-0362-7. PMID 16044244. S2CID 21096886.
- ^ Nussinov, Mark D.; Otroshchenko, Vladimir A.; Santoli, Salvatore (1997). «The emergence of the non-cellular phase of life on the fine-grained clayish particles of the early Earth’s regolith». BioSystems. 42 (2–3): 111–118. doi:10.1016/S0303-2647(96)01699-1. PMID 9184757.
- ^ Kühnlein, Alexandra; Lanzmich, Simon A.; Brun, Dieter (2 March 2021). «tRNA sequences can assemble into a replicator». eLife. 10. doi:10.7554/eLife.63431. PMC 7924937. PMID 33648631.
- ^ Noller, Harry F. (April 2012). «Evolution of protein synthesis from an RNA world». Cold Spring Harbor Perspectives in Biology. 4 (4): a003681. doi:10.1101/cshperspect.a003681. PMC 3312679. PMID 20610545.
- ^ Koonin, Eugene V. (31 May 2007). «The cosmological model of eternal inflation and the transition from chance to biological evolution in the history of life». Biology Direct. 2: 15. doi:10.1186/1745-6150-2-15. PMC 1892545. PMID 17540027.
- ^ Boone, David R.; Castenholz, Richard W.; Garrity, George M., eds. (2001). The Archaea and the Deeply Branching and Phototrophic Bacteria. Bergey’s Manual of Systematic Bacteriology. Springer. ISBN 978-0-387-21609-6. Archived from the original on 25 December 2014.[page needed]
- ^ Woese, C. R.; Fox, G. E. (1977). «Phylogenetic structure of the prokaryotic domain: the primary kingdoms». PNAS. 7 (11): 5088–5090. Bibcode:1977PNAS…74.5088W. doi:10.1073/pnas.74.11.5088. PMC 432104. PMID 270744.
- ^ Valas, R. E.; Bourne, P. E. (2011). «The origin of a derived superkingdom: how a gram-positive bacterium crossed the desert to become an archaeon». Biology Direct. 6: 16. doi:10.1186/1745-6150-6-16. PMC 3056875. PMID 21356104.
- ^ Cavalier-Smith, Thomas (2006). «Rooting the tree of life by transition analyses». Biology Direct. 1: 19. doi:10.1186/1745-6150-1-19. PMC 1586193. PMID 16834776.
- ^ «Early life liked it hot». Nature. 535 (7613): 468. 2016. doi:10.1038/535468b. S2CID 49905802.
- ^ Gogarten, Johann Peter; Deamer, David (25 November 2016). «Is LUCA a thermophilic progenote?». Nature Microbiology. 1 (12): 16229. doi:10.1038/nmicrobiol.2016.229. ISSN 2058-5276. PMID 27886195. S2CID 205428194.
- ^ Catchpole, Ryan; Forterre, Patrick (2019). «The evolution of Reverse Gyrase suggests a non-hyperthermophilic Last Universal Common Ancestor». Molecular Biology and Evolution. 36 (12): 2737–2747. doi:10.1093/molbev/msz180. PMC 6878951. PMID 31504731.
- ^ Berkemer, Sarah J.; McGlynn, Shawn E (8 August 2020). «A New Analysis of Archaea–Bacteria Domain Separation: Variable Phylogenetic Distance and the Tempo of Early Evolution». Molecular Biology and Evolution. 37 (8): 2332–2340. doi:10.1093/molbev/msaa089. PMC 7403611. PMID 32316034.
- ^ Hoffmann, Geoffrey W. (25 June 1974). «On the origin of the genetic code and the stability of the translation apparatus». Journal of Molecular Biology. 86 (2): 349–362. doi:10.1016/0022-2836(74)90024-2. PMID 4414916.
- ^ Orgel, Leslie E. (April 1963). «The Maintenance of the Accuracy of Protein Synthesis and its Relevance to Ageing». PNAS. 49 (4): 517–521. Bibcode:1963PNAS…49..517O. doi:10.1073/pnas.49.4.517. PMC 299893. PMID 13940312.
- ^ Hoffmann, Geoffrey W. (October 1975). «The Stochastic Theory of the Origin of the Genetic Code». Annual Review of Physical Chemistry. 26: 123–144. Bibcode:1975ARPC…26..123H. doi:10.1146/annurev.pc.26.100175.001011.
- ^ Brasier, M. D. (2012). Secret Chambers: The Inside Story of Cells and Complex Life. Oxford University Press. p. 298.
- ^ Ward, Peter & Kirschvink, Joe, op cit, p. 42
- ^ a b Colín-García, M.; Heredia, A.; Cordero, G.; et al. (2016). «Hydrothermal vents and prebiotic chemistry: a review». Boletín de la Sociedad Geológica Mexicana. 68 (3): 599–620. doi:10.18268/BSGM2016v68n3a13. Archived from the original on 18 August 2017.
- ^ Schirber, Michael (24 June 2014). «Hydrothermal Vents Could Explain Chemical Precursors to Life». NASA Astrobiology: Life in the Universe. NASA. Archived from the original on 29 November 2014. Retrieved 19 June 2015.
- ^ a b Martin, William; Russell, Michael J. (29 January 2003). «On the origins of cells: a hypothesis for the evolutionary transitions from abiotic geochemistry to chemoautotrophic prokaryotes, and from prokaryotes to nucleated cells». Philosophical Transactions of the Royal Society B. 358 (1429): 59–83, discussion 83–85. doi:10.1098/rstb.2002.1183. PMC 1693102. PMID 12594918.
- ^ a b Lane 2009
- ^ Usher, Oli (27 April 2015). «Chemistry of seabed’s hot vents could explain emergence of life» (Press release). University College London. Archived from the original on 20 June 2015. Retrieved 19 June 2015.
- ^ Roldan, Alberto; Hollingsworth, Nathan; Roffey, Anna; et al. (May 2015). «Bio-inspired CO2 conversion by iron sulfide catalysts under sustainable conditions». Chemical Communications. 51 (35): 7501–7504. doi:10.1039/C5CC02078F. PMID 25835242. Archived from the original on 20 June 2015. Retrieved 19 June 2015.
- ^ Baross, J. A.; Hoffman, S. E. (1985). «Submarine hydrothermal vents and associated gradient environments as sites for the origin and evolution of life». Origins of Life and Evolution of Biospheres. 15 (4): 327–345. Bibcode:1985OrLi…15..327B. doi:10.1007/bf01808177. S2CID 4613918.
- ^ Russell, M. J.; Hall, A. J. (1997). «The emergence of life from iron monosulphide bubbles at a submarine hydrothermal redox and pH front». Journal of the Geological Society. 154 (3): 377–402. Bibcode:1997JGSoc.154..377R. doi:10.1144/gsjgs.154.3.0377. PMID 11541234. S2CID 24792282.
- ^ Amend, J. P.; LaRowe, D. E.; McCollom, T. M.; Shock, E. L. (2013). «The energetics of organic synthesis inside and outside the cell». Philosophical Transactions of the Royal Society B. 368 (1622): 20120255. doi:10.1098/rstb.2012.0255. PMC 3685458. PMID 23754809.
- ^ Shock, E. L.; Boyd, E. S. (2015). «Geomicrobiology and microbial geochemistry:principles of geobiochemistry». Elements. 11: 389–394. doi:10.2113/gselements.11.6.395.
- ^ Martin, W.; Russell, M. J. (2007). «On the origin of biochemistry at an alkaline hydrothermal vent». Philosophical Transactions of the Royal Society B. 362 (1486): 1887–1925. doi:10.1098/rstb.2006.1881. PMC 2442388. PMID 17255002.
- ^ Weiss, Madeline C.; Sousa, Filipa L.; Mrnjavac, Natalia; Neukirchen, Sinje; Roettger, Mayo; Nelson-Sathi, Shijulal; Martin, William F. (25 July 2016). «The physiology and habitat of the last universal common ancestor» (PDF). Nature Microbiology. 1 (9): 16116. doi:10.1038/nmicrobiol.2016.116. PMID 27562259. S2CID 2997255.
- ^ Lane, Nick; Martin, William F. (21 December 2012). «The Origin of Membrane Bioenergetics». Cell. 151 (7): 1406–1416. doi:10.1016/j.cell.2012.11.050. ISSN 0092-8674. PMID 23260134. S2CID 15028935.
- ^ Baaske, Philipp; Weinert, Franz M.; Duhr, Stefan; Lemke, Kono H.; Russell, Michael J.; Braun, Dieter (29 May 2007). «Extreme accumulation of nucleotides in simulated hydrothermal pore systems». Proceedings of the National Academy of Sciences. 104 (22): 9346–9351. doi:10.1073/pnas.0609592104. ISSN 0027-8424. PMC 1890497. PMID 17494767.
- ^ a b c Mulkidjanian, Armen Y.; Bychkov, Andrew Yu.; Dibrova, Daria V.; Galperin, Michael Y.; Koonin, Eugene V. (3 April 2012). «Origin of first cells at terrestrial, anoxic geothermal fields». Proceedings of the National Academy of Sciences. 109 (14): E821-30. Bibcode:2012PNAS..109E.821M. doi:10.1073/pnas.1117774109. PMC 3325685. PMID 22331915.
- ^ Chandru, Kuhan; Guttenberg, Nicholas; Giri, Chaitanya; et al. (31 May 2018). «Simple prebiotic synthesis of high diversity dynamic combinatorial polyester libraries». Communications Chemistry. 1 (1). doi:10.1038/s42004-018-0031-1.
- ^ Forsythe, Jay G.; Yu, Sheng-Sheng; Mamajanov, Irena; et al. (17 August 2015). «Ester-Mediated Amide Bond Formation Driven by Wet–Dry Cycles: A Possible Path to Polypeptides on the Prebiotic Earth». Angewandte Chemie International Edition in English. 54 (34): 9871–9875. doi:10.1002/anie.201503792. PMC 4678426. PMID 26201989.
- ^ Patel, Bhavesh H.; Percivalle, Claudia; Ritson, Dougal J.; Duffy, Colm. D.; Sutherland, John D. (16 March 2015). «Common origins of RNA, protein and lipid precursors in a cyanosulfidic protometabolism». Nature Chemistry. 7 (4): 301–307. Bibcode:2015NatCh…7..301P. doi:10.1038/nchem.2202. ISSN 1755-4330. PMC 4568310. PMID 25803468.
- ^ Deamer, David (10 February 2021). «Where Did Life Begin? Testing Ideas in Prebiotic Analogue Conditions». Life. 11 (2): 134. doi:10.3390/life11020134. PMC 7916457. PMID 33578711.
- ^ Cairns-Smith, Graham (2 September 1982). Genetic Takeover and the Mineral Origins of Life. Cambridge: Cambridge University Press. ISBN 0-521-23312-7. OCLC 7875600.
- ^ Dawkins 1996, pp. 148–161
- ^ Huang, Wenhua; Ferris, James P. (12 July 2006). «One-Step, Regioselective Synthesis of up to 50-mers of RNA Oligomers by Montmorillonite Catalysis». Journal of the American Chemical Society. 128 (27): 8914–8919. doi:10.1021/ja061782k. PMID 16819887.
- ^ Subramaniam, Anand Bala; Wan, Jiandi; Gopinath, Arvind; Stone, Howard A. (2011). «Semi-permeable vesicles composed of natural clay» (PDF). Soft Matter. 7 (6): 2600–2612. arXiv:1011.4711. Bibcode:2011SMat….7.2600S. doi:10.1039/c0sm01354d. S2CID 52253528.
- ^ Hartman, Hyman (1998). «Photosynthesis and the Origin of Life». Origins of Life and Evolution of Biospheres. 28 (4–6): 515–521. Bibcode:1998OLEB…28..515H. doi:10.1023/A:1006548904157. PMID 11536891. S2CID 2464.
- ^ Yue-Ching Ho, Eugene (July–September 1990). «Evolutionary Epistemology and Sir Karl Popper’s Latest Intellectual Interest: A First-Hand Report». Intellectus. 15: 1–3. OCLC 26878740. Archived from the original on 11 March 2012.
- ^ Popper, Karl R. (29 March 1990). «Pyrite and the origin of life». Nature. 344 (6265): 387. Bibcode:1990Natur.344..387P. doi:10.1038/344387a0. S2CID 4322774.
- ^ a b Keller, Markus A.; Turchyn, Alexandra V.; Ralser, Markus (25 March 2014). «Non-enzymatic glycolysis and pentose phosphate pathway-like reactions in a plausible Archean ocean». Molecular Systems Biology. 10 (725): 725. doi:10.1002/msb.20145228. PMC 4023395. PMID 24771084.
- ^ Huber, Claudia; Wächtershäuser, Günter (31 July 1998). «Peptides by Activation of Amino Acids with CO on (Ni,Fe)S Surfaces: Implications for the Origin of Life». Science. 281 (5377): 670–672. Bibcode:1998Sci…281..670H. doi:10.1126/science.281.5377.670. PMID 9685253.
- ^ Adamala, Katarzyna; Szostak, Jack W. (29 November 2013). «Nonenzymatic Template-Directed RNA Synthesis Inside Model Protocells». Science. 342 (6162): 1098–1100. Bibcode:2013Sci…342.1098A. doi:10.1126/science.1241888. PMC 4104020. PMID 24288333.
- ^ Musser, George (23 September 2011). «How Life Arose on Earth, and How a Singularity Might Bring It Down». Observations (Blog). Archived from the original on 17 June 2015. Retrieved 17 June 2015.
- ^ Carroll, Sean (10 March 2010). «Free Energy and the Meaning of Life». Cosmic Variance (Blog). Discover. Archived from the original on 14 July 2015. Retrieved 17 June 2015.
- ^ England, Jeremy L. (28 September 2013). «Statistical physics of self-replication» (PDF). Journal of Chemical Physics. 139 (12): 121923. arXiv:1209.1179. Bibcode:2013JChPh.139l1923E. doi:10.1063/1.4818538. hdl:1721.1/90392. PMID 24089735. S2CID 478964. Archived (PDF) from the original on 4 June 2015.
- ^ Fox, Ronald F. (December 1993). «Review of Stuart Kauffman, The Origins of Order: Self-Organization and Selection in Evolution». Biophysical Journal. 65 (6): 2698–2699. Bibcode:1993BpJ….65.2698F. doi:10.1016/s0006-3495(93)81321-3. PMC 1226010.
- ^ Orgel, Leslie E. (7 November 2000). «Self-organizing biochemical cycles». PNAS. 97 (23): 12503–12507. Bibcode:2000PNAS…9712503O. doi:10.1073/pnas.220406697. PMC 18793. PMID 11058157.
- ^ Chandru, Kuhan; Gilbert, Alexis; Butch, Christopher; Aono, Masashi; Cleaves, Henderson James II (21 July 2016). «The Abiotic Chemistry of Thiolated Acetate Derivatives and the Origin of Life». Scientific Reports. 6 (29883): 29883. Bibcode:2016NatSR…629883C. doi:10.1038/srep29883. PMC 4956751. PMID 27443234.
- ^ Vallee, Yannick; Shalayel, Ibrahim; Ly, Kieu-Dung; Rao, K. V. Raghavendra; Paëpe, Gael De; Märker, Katharina; Milet, Anne (8 November 2017). «At the very beginning of life on Earth: the thiol-rich peptide (TRP) world hypothesis». The International Journal of Developmental Biology. 61 (8–9): 471–478. doi:10.1387/ijdb.170028yv. PMID 29139533.
- ^ a b Mulkidjanian, Armen Y. (24 August 2009). «On the origin of life in the zinc world: 1. Photosynthesizing, porous edifices built of hydrothermally precipitated zinc sulfide as cradles of life on Earth». Biology Direct. 4: 26. doi:10.1186/1745-6150-4-26. PMC 3152778. PMID 19703272.
- ^ Wächtershäuser, Günter (December 1988). «Before Enzymes and Templates: Theory of Surface Metabolism». Microbiological Reviews. 52 (4): 452–484. doi:10.1128/MMBR.52.4.452-484.1988. PMC 373159. PMID 3070320.
- ^ Mulkidjanian, Armen Y.; Galperin, Michael Y. (24 August 2009). «On the origin of life in the zinc world. 2. Validation of the hypothesis on the photosynthesizing zinc sulfide edifices as cradles of life on Earth». Biology Direct. 4: 27. doi:10.1186/1745-6150-4-27. PMC 2749021. PMID 19703275.
- ^ Macallum, A. B. (1 April 1926). «The Paleochemistry of the body fluids and tissues». Physiological Reviews. 6 (2): 316–357. doi:10.1152/physrev.1926.6.2.316.
- ^ Mulkidjanian, Armen Y.; Bychkov, Andrew Yu.; Dibrova, Daria V.; et al. (3 April 2012). «Origin of first cells at terrestrial, anoxic geothermal fields». PNAS. 109 (14): E821–E830. Bibcode:2012PNAS..109E.821M. doi:10.1073/pnas.1117774109. PMC 3325685. PMID 22331915.
- ^ See also Lankenau 2011, pp. 225–286, interconnecting the «Two RNA worlds» concept, and Davidovich, Chen; Belousoff, Matthew; Bashan, Anat; Yonath, Ada (September 2009). «The evolving ribosome: from non-coded peptide bond formation to sophisticated translation machinery». Research in Microbiology. 160 (7): 487–492. doi:10.1016/j.resmic.2009.07.004. PMID 19619641.
- ^ a b Plasson, Raphaël; Kondepudi, Dilip K.; Bersini, Hugues; et al. (August 2007). «Emergence of homochirality in far-from-equilibrium systems: Mechanisms and role in prebiotic chemistry». Chirality. 19 (8): 589–600. doi:10.1002/chir.20440. PMID 17559107. «Special Issue: Proceedings from the Eighteenth International Symposium on Chirality (ISCD-18), Busan, Korea, 2006»
- ^ Chaichian, Rojas & Tureanu 2014, pp. 353–364
- ^ Jafarpour, Farshid; Biancalani, Tommaso; Goldenfeld, Nigel (2017). «Noise-induced symmetry breaking far from equilibrium and the emergence of biological homochirality» (PDF). Physical Review E. 95 (3): 032407. Bibcode:2017PhRvE..95c2407J. doi:10.1103/PhysRevE.95.032407. PMID 28415353.
- ^ Jafarpour, Farshid; Biancalani, Tommaso; Goldenfeld, Nigel (2015). «Noise-induced mechanism for biological homochirality of early life self-replicators». Physical Review Letters. 115 (15): 158101. arXiv:1507.00044. Bibcode:2015PhRvL.115o8101J. doi:10.1103/PhysRevLett.115.158101. PMID 26550754. S2CID 9775791.
- ^ Frank, F.C. (1953). «On spontaneous asymmetric synthesis». Biochimica et Biophysica Acta. 11 (4): 459–463. doi:10.1016/0006-3002(53)90082-1. PMID 13105666.
- ^ Clark, Stuart (July–August 1999). «Polarized Starlight and the Handedness of Life». American Scientist. 87 (4): 336. Bibcode:1999AmSci..87..336C. doi:10.1511/1999.4.336.
- ^ Shibata, Takanori; Morioka, Hiroshi; Hayase, Tadakatsu; et al. (17 January 1996). «Highly Enantioselective Catalytic Asymmetric Automultiplication of Chiral Pyrimidyl Alcohol». Journal of the American Chemical Society. 118 (2): 471–472. doi:10.1021/ja953066g.
- ^ Soai, Kenso; Sato, Itaru; Shibata, Takanori (2001). «Asymmetric autocatalysis and the origin of chiral homogeneity in organic compounds». The Chemical Record. 1 (4): 321–332. doi:10.1002/tcr.1017. PMID 11893072.
- ^ Hazen 2005, p. 184
- ^ Meierhenrich, Uwe (2008). Amino acids and the asymmetry of life caught in the act of formation. Berlin: Springer. pp. 76–79. ISBN 978-3540768869.
- ^ Mullen, Leslie (5 September 2005). «Building Life from Star-Stuff». Astrobiology Magazine. Archived from the original on 14 July 2015.
General and cited sources[edit]
- Altermann, Wladyslaw (2009). «Introduction: A Roadmap to Fata Morgana?». In Seckbach, Joseph; Walsh, Maud (eds.). From Fossils to Astrobiology: Records of Life on Earth and the Search for Extraterrestrial Biosignatures. Cellular Origin, Life in Extreme Habitats and Astrobiology. Vol. 12. Dordrecht, the Netherlands; London: Springer. ISBN 978-1-4020-8836-0.
- Bada, Jeffrey L.; Lazcano, Antonio (2009). «The Origin of Life». In Ruse, Michael; Travis, Joseph (eds.). Evolution: The First Four Billion Years. Foreword by Edward O. Wilson. Cambridge: Belknap Press of Harvard University Press. ISBN 978-0-674-03175-3. OCLC 225874308.
- Barton, Nicholas H.; Briggs, Derek E.G.; Eisen, Jonathan A.; et al. (2007). Evolution. Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press. ISBN 978-0-87969-684-9. OCLC 86090399.
- Bastian, H. Charlton (1871). The Modes of Origin of Lowest Organisms. London; New York: Macmillan and Company. OCLC 42959303. Retrieved 6 June 2015.
- Bernal, J. D. (1951). The Physical Basis of Life. London: Routledge & Kegan Paul.
- Bernal, J. D. (1960). «The Problem of Stages in Biopoesis». In Florkin, M. (ed.). Aspects of the Origin of Life. International Series of Monographs on Pure and Applied Biology. Oxford, UK; New York: Pergamon Press. ISBN 978-1-4831-3587-8.
- Bernal, J. D. (1967) [Reprinted work by A.I. Oparin originally published 1924; Moscow: The Moscow Worker]. The Origin of Life. The Weidenfeld and Nicolson Natural History. Translation of Oparin by Ann Synge. London: Weidenfeld & Nicolson.
- Bock, Gregory R.; Goode, Jamie A., eds. (1996). Evolution of Hydrothermal Ecosystems on Earth (and Mars?). Ciba Foundation Symposium. Vol. 202. Chichester, UK; New York: John Wiley & Sons. ISBN 978-0-471-96509-1.
- Bondeson, Jan (1999). The Feejee Mermaid and Other Essays in Natural and Unnatural History. Ithaca, NY: Cornell University Press. ISBN 978-0-8014-3609-3.
- Bryson, Bill (2004). A Short History of Nearly Everything. London: Black Swan. ISBN 978-0-552-99704-1. OCLC 55589795.
- Calvin, Melvin (1969). Chemical Evolution: Molecular Evolution Towards the Origin of Living Systems on the Earth and Elsewhere. Oxford, UK: Clarendon Press. ISBN 978-0-19-855342-7. OCLC 25220.
- Chaichian, Masud; Rojas, Hugo Perez; Tureanu, Anca (2014). «Physics and Life». Basic Concepts in Physics: From the Cosmos to Quarks. Undergraduate Lecture Notes in Physics. Berlin; Heidelberg: Springer Berlin Heidelberg. doi:10.1007/978-3-642-19598-3_12. ISBN 978-3-642-19597-6. OCLC 900189038. S2CID 115247432.
- Chang, Thomas Ming Swi (2007). Artificial Cells: Biotechnology, Nanomedicine, Regenerative Medicine, Blood Substitutes, Bioencapsulation, and Cell/Stem Cell Therapy. Regenerative Medicine, Artificial Cells and Nanomedicine. Vol. 1. Hackensack, New Jersey: World Scientific. ISBN 978-981-270-576-1. OCLC 173522612.
- Dalrymple, G. Brent (2001). «The age of the Earth in the twentieth century: a problem (mostly) solved». In Lewis, C.L.E.; Knell, S.J. (eds.). The Age of the Earth: from 4004 BC to AD 2002. Geological Society of London, Special Publications. Geological Society Special Publication. Vol. 190. London: Geological Society of London. pp. 205–221. Bibcode:2001GSLSP.190..205D. doi:10.1144/gsl.sp.2001.190.01.14. ISBN 978-1-86239-093-5. OCLC 48570033. S2CID 130092094.
- Darwin, Charles (1887). Darwin, Francis (ed.). The Life and Letters of Charles Darwin, Including an Autobiographical Chapter. Vol. 3 (3rd ed.). London: John Murray. OCLC 834491774.
- Davies, Paul (1999). The Fifth Miracle: The Search for the Origin of Life. London: Penguin Books. ISBN 978-0-14-028226-9.
- Dawkins, Richard (1996). The Blind Watchmaker (Reissue with a new introduction ed.). New York: W.W. Norton & Company. ISBN 978-0-393-31570-7. OCLC 35648431.
- Dobell, Clifford (1960) [Originally published 1932; New York: Harcourt, Brace & Company]. Antony van Leeuwenhoek and His ‘Little Animals’. New York: Dover Publications.
- Dyson, Freeman (1999). Origins of Life (Revised ed.). Cambridge, UK; New York: Cambridge University Press. ISBN 978-0-521-62668-2.
- Fesenkov, V. G. (1959). «Some Considerations about the Primaeval State of the Earth». In Oparin, A.I.; et al. (eds.). The Origin of Life on the Earth. I.U.B. Symposium Series. Vol. 1. Edited for the International Union of Biochemistry by Frank Clark and R.L.M. Synge (English-French-German ed.). London; New York: Pergamon Press. ISBN 978-1-4832-2240-0. International Symposium on the Origin of Life on the Earth (held at Moscow, 19–24 August 1957)
- Hazen, Robert M. (2005). Genesis: The Scientific Quest for Life’s Origin. Washington, DC: Joseph Henry Press. ISBN 978-0-309-09432-0. OCLC 60321860.
- Huxley, Thomas Henry (1968) [1897]. «VIII Biogenesis and Abiogenesis [1870]». Discourses, Biological and Geological. Collected Essays. Vol. VIII (Reprint ed.). New York: Greenwood Press. OCLC 476737627.
- Klyce, Brig (22 January 2001). Kingsley, Stuart A.; Bhathal, Ragbir (eds.). Panspermia Asks New Questions. The Search for Extraterrestrial Intelligence (SETI) in the Optical Spectrum III. Vol. 4273. Bellingham, WA: SPIE. doi:10.1117/12.435366. ISBN 0-8194-3951-7. Retrieved 9 June 2015. Proceedings of the SPIE held at San Jose, California, 22–24 January 2001
- Lane, Nick (2009). Life Ascending: The 10 Great Inventions of Evolution (1st American ed.). New York: W.W. Norton & Company. ISBN 978-0-393-06596-1. OCLC 286488326.
- Lankenau, Dirk-Henner (2011). «Two RNA Worlds: Toward the Origin of Replication, Genes, Recombination and Repair». In Egel, Richard; Lankenau, Dirk-Henner; Mulkidjanian, Armen Y. (eds.). Origins of Life: The Primal Self-Organization. Heidelberg: Springer. doi:10.1007/978-3-642-21625-1. ISBN 978-3-642-21624-4. OCLC 733245537.
- Lennox, James G. (2001). Aristotle’s Philosophy of Biology: Studies in the Origins of Life Science. Cambridge Studies in Philosophy and Biology. Cambridge, UK; New York: Cambridge University Press. ISBN 978-0-521-65976-5.
- Michod, Richard E. (1999). Darwinian Dynamics: Evolutionary Transitions in Fitness and Individuality. Princeton, NJ: Princeton University Press. ISBN 978-0-691-02699-2. OCLC 38948118.
- Oparin, A.I. (1953) [Originally published 1938; New York: The Macmillan Company]. The Origin of Life. Translation and new introduction by Sergius Morgulis (2nd ed.). Mineola, NY: Dover Publications. ISBN 978-0-486-49522-4.
- Raven, Peter H.; Johnson, George B. (2002). Biology (6th ed.). Boston: McGraw-Hill. ISBN 978-0-07-112261-0. OCLC 45806501.
- Ross, Alexander (1652). Arcana Microcosmi. Vol. II. London. OCLC 614453394.
- Sheldon, Robert B. (22 September 2005). «Historical Development of the Distinction between Bio- and Abiogenesis» (PDF). In Hoover, Richard B.; Levin, Gilbert V.; Rozanov, Alexei Y.; Gladstone, G. Randall (eds.). Astrobiology and Planetary Missions. Astrobiology and Planetary Missions. Vol. 5906. Bellingham, WA: SPIE. pp. 59061I. doi:10.1117/12.663480. ISBN 978-0-8194-5911-4. Proceedings of the SPIE held at San Diego, California, 31 July–2 August 2005
- Tyndall, John (1905) [Originally published 1871; London; New York: Longmans, Green & Co.; D. Appleton and Company]. Fragments of Science. Vol. 2 (6th ed.). New York: P.F. Collier & Sons. OCLC 726998155.
- Voet, Donald; Voet, Judith G. (2004). Biochemistry. Vol. 1 (3rd ed.). New York: John Wiley & Sons. ISBN 978-0-471-19350-0.
- Yarus, Michael (2010). Life from an RNA World: The Ancestor Within. Cambridge, Massachusetts: Harvard University Press. p. 287. ISBN 978-0-674-05075-4.
External links[edit]
- Making headway with the mysteries of life’s origins – Adam Mann (PNAS; 14 April 2021)
- How life began on Earth – Marcia Malory (Earth Facts; 2015)
- The Origins of Life – Richard Dawkins et al. (BBC Radio; 2004)
- Life in the Universe – Essay by Stephen Hawking (1996)
«Происхождение жизни» перенаправляется сюда. О ненаучных взглядах на происхождение жизни см. Миф о сотворении мира.
В эволюционная биология, абиогенез, или неофициально происхождение жизни (OoL),[3][4][5][а] это естественный процесс, посредством которого жизнь возник из неживой материи, такой как простой органические соединения.[6][4][7][8] Хотя детали этого процесса до сих пор неизвестны, преобладающая научная гипотеза состоит в том, что переход от неживых к живым существам был не единичным событием, а эволюционным процессом все возрастающей сложности, который включал молекулярные самовоспроизведение, самосборка, автокатализ, и появление клеточные мембраны.[9][10][11] Хотя возникновение абиогенеза не вызывает споров среди ученых, его возможные механизмы плохо изучены. Есть несколько принципов и гипотез для как мог произойти абиогенез.[12]
Изучение абиогенеза направлено на определение того, как до жизни химические реакции дали начало жизни в условиях, разительно отличных от тех, которые существуют на Земле сегодня.[13] В основном он использует инструменты из биология, химия, и геофизика,[14] с более поздними подходами, пытающимися синтезировать все три:[15] более конкретно, астробиология, биохимия, биофизика, геохимия, молекулярная биология, океанография и палеонтология. Жизнь функционирует через специализированную химию углерод и вода и основывается в основном на четырех ключевых группах химических веществ: липиды (клеточные мембраны), углеводы (сахара, целлюлоза), аминокислоты (метаболизм белков) и нуклеиновые кислоты (ДНК и РНК). Любая успешная теория абиогенеза должна объяснить происхождение и взаимодействие этих классов молекул.[16] Многие подходы к абиогенезу исследуют, как самовоспроизводящийся молекулы или их компоненты. Исследователи обычно считают, что текущая жизнь происходит от Мир РНК,[17] хотя другие самовоспроизводящиеся молекулы могли предшествовать РНК.[18][19]

Эксперимент Миллера – Юри Синтез небольших органических молекул в смеси простых газов, которые помещаются в температурный градиент путем одновременного нагрева (слева) и охлаждения (справа) смеси, смеси, которая также подвержена электрическим разрядам
Классика 1952 года Эксперимент Миллера – Юри и аналогичные исследования показали, что большинство аминокислот, химических составляющих белки используется во всех живых организмах, может быть синтезирован из неорганические соединения в условиях, предназначенных для воспроизведения тех из ранняя земля. Ученые предложили различные внешние источники энергии, которые могли вызвать эти реакции, в том числе: молния и радиация. Другие подходы (гипотезы «сначала метаболизм») сосредоточены на понимании того, как катализ в химических системах на ранней Земле могли обеспечить молекулы-предшественники необходимо для самовоспроизведения.[20]
Альтернатива гипотеза панспермии[21] предполагает, что микроскопическая жизнь возник за пределами Земли неизвестными механизмами и распространился на раннюю Землю на космическая пыль[22] и метеороиды.[23] Известно, что комплекс Органические молекулы происходят в Солнечная система И в межзвездное пространство, и эти молекулы могли обеспечить исходный материал для развития жизни на Земле.[24][25][26][27]
Земля остается единственным местом в мире вселенная известно, что укрывает жизнь,[28][29] и ископаемые свидетельства с Земли информирует большинство исследований абиогенеза. В возраст Земли составляет 4,54 Гр (миллиард год);[30][31][32] самое раннее неоспоримое свидетельство существования жизни на Земле датируется по крайней мере 3,5 Гя (Гр назад),[33][34][35] и, возможно, уже в Эоархейский Эра (3,6–4,0 Гя). В 2017 году ученые нашли возможные доказательства ранней жизни на земле в 3,48 Гё (Гр) гейзерит и другие месторождения полезных ископаемых (часто встречаются вокруг горячие источники и гейзеры ) раскрытые в Пилбара Кратон из Западная Австралия.[36][37][38][39] Однако ряд открытий предполагает, что жизнь могла появиться на Земле еще раньше. По состоянию на 2017 год, микрофоссилий (окаменелые микроорганизмы ) в осадки гидротермальных источников от 3,77 до 4,28 Гя в породах в Квебек может содержать старейшие летописи жизни на Земле, предполагая, что жизнь началась вскоре после образование океана 4,4 Гя вовремя Hadean Eon.[1][2][40][41][42]
Стратегия НАСА по абиогенезу утверждает, что необходимо идентифицировать взаимодействия, промежуточные структуры и функции, источники энергии и факторы окружающей среды, которые способствовали разнообразию, отбору и репликации эволюционирующих макромолекулярных систем.[43] Акцент должен продолжаться на нанесении на карту химического ландшафта потенциальных первичных информационных полимеры. Появление полимеров, которые могут реплицироваться, хранить генетическую информацию и проявлять свойства, подлежащие отбору, вероятно, было решающим шагом в появление пребиотической химической эволюции.[43]
Термодинамика, самоорганизация и информация: Физика
Принципы термодинамики: энергия и энтропия
В древности обычно считалось, например, Эмпедоклом и Аристотелем, что жизнь особей некоторых видов и, в более общем плане, сама жизнь может начинаться с высокой температуры, то есть неявно за счет теплового цикла.[44]
Точно так же было рано осознавать, что жизнь требует потери энтропия, или беспорядок, когда молекулы организуются в живую материю. Этот Второй закон термодинамики необходимо учитывать, когда происходит самоорганизация материи до более высокой сложности. Потому что живые организмы — это машины,[45] Второй Закон применим и к жизни.
Таким образом, вопреки наивному взгляду на Второй закон, возникновение жизни и ее усложнение не противоречат этому закону: во-первых, живой организм создает порядок в некоторых местах (например, в своем живом теле или жилище) за счет увеличения энтропия в другом месте (например, производство тепла и отходов). Во-вторых, второй закон термодинамики фактически предсказывает увеличение сложности[46] и во взаимосвязях между системой и ее окружением при взаимодействии[47] — память и генетическая адаптация являются примерами таких взаимосвязей между живым организмом и окружающей его средой.
Получение бесплатной энергии
Бернал сказал об эксперименте Миллера – Юри, что
недостаточно для объяснения образования таких молекул, необходимо физико-химическое объяснение происхождения этих молекул, которое предполагает наличие подходящих источников и стоков для свободной энергии.[48]
На ранней Земле для химических реакций было доступно множество источников энергии. Например, тепло (например, от геотермальный процессы) является стандартным источником энергии для химии. Среди других примеров — солнечный свет и электрические разряды (молния).[49] Фактически, молния является вероятным источником энергии для зарождения жизни, учитывая, что только в тропиках молния ударяет около 100 миллионов раз в год.[50]
Компьютерное моделирование также предполагает, что кавитация в первичных водоемах, таких как набегающие морские волны, ручьи и океаны, потенциально может привести к синтезу биогенных соединений.[51]
Неблагоприятные реакции также могут быть вызваны очень благоприятными, как в случае химии железа и серы. Например, это, вероятно, было важно для фиксация углерода (превращение углерода из неорганической формы в органическую).[b] Фиксация углерода посредством химии железа и серы очень благоприятна и происходит при нейтральном pH и 100 ° C. Поверхности, содержащие железо-серу, которых много около гидротермальных источников, также способны производить небольшие количества аминокислот и других биологических метаболитов.[49]
Самоорганизация

Герман Хакен
Дисциплина синергетики изучает самоорганизацию физических систем. В его книге Синергетика[52] Герман Хакен указал, что разные физические системы можно рассматривать одинаково. Он приводит в качестве примеров самоорганизации несколько типов лазеров, нестабильности в гидродинамике, включая конвекцию, а также химические и биохимические колебания. В своем предисловии он упоминает происхождение жизни, но только в общих чертах:
Спонтанное образование хорошо организованных структур из микробов или даже из хаоса — одно из самых захватывающих явлений и самых сложных проблем, с которыми сталкиваются ученые. Такие явления — это опыт нашей повседневной жизни, когда мы наблюдаем рост растений и животных. Размышляя о гораздо больших временных масштабах, ученые сталкиваются с проблемами эволюции и, в конечном итоге, происхождения живого вещества. Когда мы пытаемся объяснить или понять в некотором смысле эти чрезвычайно сложные биологические явления, возникает естественный вопрос, можно ли найти процессы самоорганизации в гораздо более простых системах неодушевленного мира.
В последние годы становится все более очевидным, что существует множество примеров в физических и химических системах, где хорошо организованные пространственные, временные или пространственно-временные структуры возникают из хаотических состояний. Более того, как и в живых организмах, функционирование этих систем может поддерживаться только потоком энергии (и вещества) через них. В отличие от машин, созданных руками человека, которые предназначены для демонстрации особых структур и функций, эти структуры развиваются спонтанно — они самоорганизующийся. …[53]
Множественные диссипативные структуры
Эта теория постулирует, что отличительной чертой происхождения и эволюции жизни является микроскопическое диссипативное структурирование органические пигменты и их распространение по всей поверхности Земли.[54] Современная жизнь увеличивает производство энтропии Земли в ее солнечной среде, рассеивая ультрафиолетовый и видимый фотоны в тепло через органические пигменты в воде. Затем это тепло катализирует множество вторичных диссипативных процессов, таких как круговорот воды, океан и ветер токи, ураганы, так далее.[55][56]
Самоорганизация диссипативными структурами

Илья Пригожин 1977c
Физик XIX века Людвиг Больцманн первым осознал, что борьба за существование живых организмов велась не из-за сырья или энергия, но вместо этого имел дело с производство энтропии полученный из преобразования солнечной спектр в высокая температура этими системами.[57] Таким образом, Больцман понял, что живые системы, как и все необратимые процессы, зависели от рассеяние обобщенного химического потенциала их существования. В своей книге «Что такое жизнь» физик 20 века Эрвин Шредингер[58] подчеркнули важность глубокого понимания Больцманом необратимой термодинамической природы живых систем, предположив, что это были физика и химия, лежащие в основе происхождения и эволюции жизни.
Однако необратимые процессы и тем более живые системы не могли быть удобно проанализированы с этой точки зрения до тех пор, пока Ларс Онсагер,[59] а позже Илья Пригожин,[60] разработал элегантный математический формализм для рассмотрения «самоорганизации» материала под действием обобщенного химического потенциала. Этот формализм стал известен как классическая необратимая термодинамика, и Пригожин был удостоен награды Нобелевская премия по химии в 1977 г. «за его вклад в неравновесная термодинамика, особенно теория диссипативные структуры «. Анализ Пригожина показал, что если система были оставлены для развития под навязанным внешним потенциалом, материал мог спонтанно организовываться (снижать энтропия ) образуя то, что он назвал «диссипативными структурами», которые увеличивают диссипацию внешнего навязанного потенциала (увеличивают производство глобальной энтропии). Неравновесная термодинамика с тех пор успешно применялась для анализа живых систем, начиная с биохимического производства АТФ[61] для оптимизации бактериальных метаболических путей[62] для завершения экосистем.[63][64][65]
Современная жизнь, результат абиогенеза: биология
Определение жизни
Составлено 123 определения жизни.[66]
При обсуждении происхождения жизни фундаментальным является определение самой жизни. С этим определением несколько разошлись (хотя и следует одним и тем же основным принципам), потому что разные учебники биологии определяют жизнь по-разному. Джеймс Гулд:
Большинство словарей определяют жизнь как свойство, которое отличает живых от мертвых, и определяет мертвых как лишенный жизни. Эти необычно круглые и неудовлетворительные определения не дают нам ни малейшего представления о том, что у нас общего с простейшими и растениями. [67]
тогда как по словам Нила Кэмпбелла и Джейн Рис
Феномен, который мы называем жизнью, не поддается простому определению, состоящему из одного предложения.[68]
Это различие также можно найти в книгах о происхождении жизни. Джон Касти дает единственное предложение:
По более или менее общему мнению, в настоящее время сущность считается «живой», если она способна выполнять три основных функциональных действия: метаболизм, самовосстановление и репликацию.[69]
Дирк Шульце-Макух и Луи Ирвин, напротив, посвятили всю первую главу своей книги этой теме.[70]
Ферментация

Цикл лимонной кислоты

Общая схема химических реакций обмена веществ, в которой цикл лимонной кислоты можно распознать в виде круга чуть ниже середины рисунка.
Альберт Ленингер примерно в 1970 году заявил, что ферментация, включая гликолиз, является подходящим примитивным источником энергии для возникновения жизни.[71]
Поскольку живые организмы, вероятно, впервые возникли в атмосфере, лишенной кислорода, анаэробная ферментация является самым простым и наиболее примитивным типом биологического механизма получения энергии из молекул питательных веществ.
Ферментация включает гликолиз, который довольно неэффективно преобразует химическую энергию сахара в химическую энергию АТФ.
Хемиосмос
Окислительного фосфорилирования

Хемиосмотическое соединение митохондрий
В качестве Ферментация Когда примерно в 1970 г. был выяснен, в то время как механизм окислительного фосфорилирования не был выяснен, а некоторые споры все еще существовали, ферментация могла показаться слишком сложной для исследователей происхождения жизни в то время. Питер Митчелл с Хемиосмос однако сейчас принято считать правильным.
Даже сам Питер Митчелл предполагал, что брожение предшествовало хемиосмосу. Однако хемиосмос встречается в жизни повсеместно. Модель происхождения жизни была представлена с точки зрения хемиосмоса.[72][73]
И дыхание митохондриями, и фотосинтез в хлоропластах используют хемиосмос для генерации большей части своего АТФ.
Сегодня источник энергии для всего живого можно связать с фотосинтезом, и говорят о первичном производстве солнечным светом. Кислород, используемый для окисления восстанавливающих соединений организмами в гидротермальных источниках на дне океана, является результатом фотосинтеза на поверхности Мирового океана.
АТФ-синтаза

Пол Бойер
Механизм синтеза АТФ сложен и включает закрытую мембрану, в которой АТФ-синтаза встроен. АТФ синтезируется субъединицей F1 АТФ-синтазы посредством механизм изменения привязки обнаружен Пол Бойер. Энергия, необходимая для высвобождения сформированного прочно связанного АТФ, происходит от протонов, которые перемещаются через мембрану. Эти протоны проходят через мембрану во время дыхания или фотосинтеза.
Мир РНК

В Мир РНК Гипотеза описывает раннюю Землю с самовоспроизводящейся и каталитической РНК, но без ДНК или белков.[75] Широко признано, что современная жизнь на Земле происходит из мира РНК,[17][76][77] хотя жизнь, основанная на РНК, возможно, не была первой существующей жизнью.[18][19] Этот вывод сделан на основании множества независимых доказательств, таких как наблюдения, что РНК играет центральную роль в процессе трансляции и что малые РНК могут катализировать все химические группы и передачу информации, необходимые для жизни.[19][78] Структура рибосома был назван «дымящимся пистолетом», поскольку показал, что рибосома представляет собой рибозим с центральным ядром РНК и без боковых цепей аминокислот в пределах 18 ангстрем от активный сайт где катализируется образование пептидной связи.[18][79]
Концепция мира РНК была впервые предложена в 1962 г. Александр Рич,[80] и термин был придуман Уолтер Гилберт в 1986 г.[19][81]В марте 2020 года астроном Томонори Тотани представил статистический подход для объяснения того, как исходная активная молекула РНК могла произвольно образовываться в вселенная когда-то с Большой взрыв.[82][83]
Филогения и LUCA
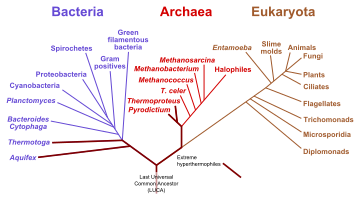
Наиболее общепринятое расположение корня древа жизни — между монофилетической областью. Бактерии и кладу, образованную Археи и Эукариоты того, что называют «традиционным деревом жизни» на основании нескольких молекулярных исследований, начиная с Карл Вёзе.[84][85]
Очень незначительное меньшинство исследований пришли к другому выводу, а именно, что корень находится в домене Bacteria, либо в типе Фирмикуты[86] или что тип Хлорофлекси базальна по отношению к кладе с архей + эукариотами и остальными бактериями, как было предложено Томас Кавалье-Смит.[87] В последнее время, Питер Уорд предложил альтернативную точку зрения, основанную на синтезе абиотической РНК, которая оказывается заключенной в капсулу и затем создает РНК. рибозим реплицирует. Предполагается, что это затем раздвоится между Dominion Ribosa (РНК жизнь ), а после потери рибозимов РНК вирусов как Domain Viorea и Dominion Terroa[требуется разъяснение ], который после создания большой клетки внутри липидной стенки, создания ДНК, состоящей из 20 аминокислот и триплетного кода, устанавливается как последний универсальный общий предок или LUCA, более ранних филогенных деревьев.[88]
В 2016 году набор из 355 штук. гены вероятно присутствует в Последний универсальный общий предок (LUCA) всех организмы живущий на Земле был идентифицирован.[89] Всего было секвенировано 6,1 миллиона генов, кодирующих прокариотические белки, из различных филогенных деревьев, что позволило идентифицировать 355 кластеров белков из 286 514 кластеров белков, которые, вероятно, были общими для LUCA. Результаты, достижения
. . . изображаем LUCA как анаэробный, CO2-фиксирующий, H2-зависимый с Дорога Вуд – Юнгдал, N2-фиксирующий и теплолюбивый. Биохимия LUCA изобилует кластерами FeS и механизмами радикальных реакций. Его кофакторы выявить зависимость от переходные металлы, флавины, S-аденозил метионин, коэнзим А, ферредоксин, молибдоптерин, Corrins и селен. Его генетический код требовал нуклеозид модификации и S-аденозилметионин-зависимые метилирования.»
Результаты отображают метаногенный клостридия в качестве базальной клады в 355 изученных филогенезах и предполагают, что LUCA населяла анаэробные гидротермальный источник обстановка в геохимически активной среде, богатой H2, CO2 и железо.[90]
Исследование в Университете Дюссельдорфа создало филогенные деревья на основе 6 миллионов генов из бактерии и археи, и идентифицировали 355 семейств белков, которые, вероятно, присутствовали в LUCA. Они были основаны на фиксации анаэробного метаболизма. углекислый газ и азот. Это предполагает, что LUCA развивалась в среде, богатой водород, двуокись углерода и утюг.[91]
Ключевые вопросы абиогенеза
Что было раньше: белок или нуклеиновые кислоты?
Возможные предшественники для эволюции синтеза белка включают механизм синтеза коротких пептидных кофакторов или формирование механизма дублирования РНК. Вероятно, что предковая рибосома полностью состояла из РНК, хотя некоторые роли с тех пор взяли на себя белки. Основные оставшиеся вопросы по этой теме включают определение силы отбора для эволюции рибосомы и определение того, как генетический код возникла.[92]
Евгений Кунин сказал,
Несмотря на значительные экспериментальные и теоретические усилия, в настоящее время не существует убедительных сценариев происхождения репликации и трансляции, ключевых процессов, которые вместе составляют ядро биологических систем и очевидную предпосылку биологической эволюции. Концепция мира РНК может предложить лучший шанс для решения этой головоломки, но пока не может адекватно объяснить появление эффективной репликазы РНК или системы трансляции. MWO [«много миров в одном»] версия космологической модели вечная инфляция может предложить выход из этой головоломки, потому что в бесконечном мультивселенная с конечным числом различных макроскопических историй (каждая повторяется бесконечное число раз) случайное появление даже очень сложных систем не только возможно, но и неизбежно.[93]
Появление генетического кода
Видеть: Генетический код.
Ошибка в переводе катастрофа
Хоффманн показал, что ранний механизм перевода, подверженный ошибкам, может быть устойчивым к катастрофическим ошибкам того типа, который считался проблематичным для происхождения жизни и был известен как «парадокс Оргеля».[94][95][96]
Гомохиральность
Гомохиральность относится к геометрической однородности некоторых материалов, состоящих из хиральный единицы. Киральный относится к несуперпространственным трехмерным формам, которые являются зеркальным отображением друг друга, как левая и правая руки. Живые организмы используют молекулы с одинаковой хиральностью («хиральностью»): почти без исключений,[97] аминокислоты являются левыми, а нуклеотиды и сахара правши. Хиральные молекулы могут быть синтезированы, но в отсутствие хирального источника или хирального катализатор, они образованы в смеси 50/50 обоих энантиомеры (так называемая рацемическая смесь). Известные механизмы производства нерацемических смесей из рацемических исходных материалов включают: асимметричные физические законы, такие как электрослабое взаимодействие; асимметричная среда, например, вызванная циркулярно поляризованный свет, кристаллы кварца, или вращение Земли, статистические колебания во время рацемического синтеза,[98] и спонтанное нарушение симметрии.[99][100][101]
После установления хиральности будет выбрана.[102] Небольшой перекос (энантиомерный избыток ) в популяции может быть увеличена до большого асимметричный автокатализ, например, в Соаи реакция.[103] В асимметричном автокатализе катализатор представляет собой хиральную молекулу, что означает, что хиральная молекула катализирует собственное производство. Первоначальный избыток энантиомеров, который может быть получен поляризованным светом, затем позволяет более распространенному энантиомеру вытеснить другой.[104]
Кларк предположил, что гомохиральность могла начаться в космическом пространстве, поскольку исследования аминокислот на Метеорит Мерчисон показало, что L-аланин более чем в два раза чаще, чем его форма D, и L-глутаминовая кислота был более чем в три раза распространен, чем его коллега D. Различные хиральные кристаллические поверхности также могут действовать как места для возможной концентрации и сборки хиральных мономерных звеньев в макромолекулы.[105][106] Соединения, обнаруженные на метеоритах, предполагают, что хиральность жизни происходит из абиогенного синтеза, поскольку аминокислоты из метеоритов демонстрируют левое смещение, тогда как сахара демонстрируют преимущественно правое смещение, такое же, как у живых организмов.[107]
Ранняя Вселенная и Земля: астрономия и геология
Ранняя вселенная с первыми звездами
-13 —
–
-12 —
–
-11 —
–
-10 —
–
-9 —
–
-8 —
–
-7 —
–
-6 —
–
-5 —
–
-4 —
–
-3 —
–
-2 —
–
-1 —
–
0 —
-4500 —
–
-4000 —
–
-3500 —
–
-3000 —
–
-2500 —
–
-2000 —
–
-1500 —
–
-1000 —
–
-500 —
–
0 —
Вскоре после Большой взрыв, который произошел примерно в 14 Гйа, единственными химическими элементами, присутствующими во Вселенной, были водород, гелий и литий, три самых легких атома в периодической таблице. Эти элементы постепенно сошлись, чтобы сформировать звезды. Эти ранние звезды были массивными и недолговечными, производя более тяжелые элементы через звездный нуклеосинтез. Углерод, в настоящее время четвертый по распространенности химический элемент во вселенной (после водород, гелий и кислород ), сформировалась в основном в белые карликовые звезды, особенно те, которые больше двух солнечных масс.[108][109]
Когда эти звезды достигли конца своего жизненные циклы, они выбросили эти более тяжелые элементы, в том числе углерод и кислород, по всей Вселенной. Эти более тяжелые элементы позволили сформировать новые объекты, включая скалистые планеты и другие тела.[110]
Возникновение Солнечной системы
Согласно небулярная гипотеза, формирование и эволюция Солнечная система началась 4.6 Гя с гравитационный коллапс небольшой части гиганта молекулярное облако.[111] Большая часть разрушающейся массы собралась в центре, образуя солнце, а остальные расплющились в протопланетный диск из которых планеты, луны, астероиды, и другие небольшие тела Солнечной системы сформирован.
Возникновение Земли
Земля, образованная 4,5 Гя, поначалу была неприветливой для любых живых организмов. На основании многочисленных наблюдений и исследований геологическая шкала времени, то Hadean Считается, что Земля пережила вторичная атмосфера, сформированный через дегазация пород, накопившихся из планетезимальный ударники. Сначала считалось, что Земля атмосфера состоял из водородных соединений -метан, аммиак и Водяной пар — и жизнь началась при таком сокращение условия, способствующие образованию органических молекул. Согласно более поздним моделям, предложенным при изучении древних минералов, атмосфера в поздний хадейский период состояла в основном из водяного пара. азот и углекислый газ, с меньшим количеством монооксид углерода, водород, и сера соединения.[112] Во время своего формирования Земля потеряла значительную часть своей первоначальной массы, при этом осталось ядро из более тяжелых скалистых элементов протопланетного диска.[113] Как следствие, Земле не хватало сила тяжести удерживать любой молекулярный водород в своей атмосфере и быстро терять его в течение периода Хадея вместе с основной массой исходных инертных газов. Считается, что раствор углекислого газа в воде слегка изменил море кислый, давая им pH около 5,5.[114] Атмосфера в то время характеризовалась как «гигантская производительная химическая лаборатория под открытым небом».[49] Возможно, это было похоже на смесь газов, выпущенных сегодня вулканами, которые все еще поддерживают некоторую абиотическую химию.[49]
Возникновение океана
Океаны можно иметь появился первым в Хадейском эоне, через 200 млн. лет после образования Земли, в горячей, 100 ° C, восстанавливающей среде, и pH около 5,8 быстро повысился до нейтрального.[115] Этот сценарий нашел подтверждение в датировке 4,404 Gyo. циркон кристаллы из метаморфизованных кварцит из Гора Нарраер в Западной Австралии Джек Хиллз из Пилбара, которые свидетельствуют о том, что океаны и Континентальный разлом существовало в пределах 150Ма формирования Земли.[116] Несмотря на вероятный рост вулканизма и существование множества более мелких тектонический «тромбоциты», было высказано предположение, что между 4,4-4,3 Гл. Земля была водным миром с небольшой континентальной корой, если вообще имела ее, чрезвычайно бурный атмосфера и гидросфера при условии интенсивного ультрафиолетовый (УФ) свет от Т Тельца, стадия Солнца, космическое излучение и продолжил болид ударов.[117]
Поздняя тяжелая бомбардировка
Окружающая среда Хадея была бы очень опасной для современной жизни. Частых столкновений с крупными объектами до 500 км в диаметре было бы достаточно, чтобы стерилизовать планету и испарить океаны в течение нескольких месяцев после столкновения, при этом горячий пар, смешанный с паром горных пород, превратился в высокогорные облака, которые полностью покрыли бы планету. Через несколько месяцев высота этих облаков начала бы уменьшаться, но основание облака все еще оставалось бы приподнятым в течение примерно следующей тысячи лет. После этого на небольшой высоте пошел бы дождь. Еще две тысячи лет дожди медленно опустили бы облака, вернув океаны на их первоначальную глубину только через 3000 лет после столкновения.[118]
Традиционно считалось, что в период с 4.28 г.[1][2] и 3,8 Гя — изменения орбит планеты-гиганты мог вызвать сильная бомбардировка астероидами и кометы[119] что испортило Луна и другие внутренние планеты (Меркурий, Марс, и предположительно Земля и Венера ). Это, вероятно, неоднократно стерилизовало бы планету, если бы до этого времени появилась жизнь.[49] В геологическом отношении Хадейская Земля была бы намного активнее, чем когда-либо в своей истории. Исследования метеориты предполагает, что радиоактивные изотопы Такие как алюминий-26 с период полураспада 7,17 тыс. лет назад, и калий-40 с периодом полураспада 1,25 Гр, изотопы в основном производятся в сверхновые, были гораздо чаще.[120] Внутреннее отопление в результате гравитационная сортировка между основной и мантия вызвало бы много мантийная конвекция, с вероятным результатом гораздо большего количества меньших и более активных тектонических плит, чем существует сейчас.
Промежутки времени между такими разрушительными экологическими событиями дают временные окна для возможного происхождения жизни в ранних средах. Если бы глубоководная гидротермальная обстановка была местом зарождения жизни, то абиогенез мог произойти уже в 4,0-4,2 млрд лет назад. Если бы это место находилось на поверхности Земли, абиогенез мог произойти только между 3,7-4,0 Гя.[121]
Оценки производства органических веществ из этих источников показывают, что Поздняя тяжелая бомбардировка до 3,5 млрд лет назад в ранней атмосфере стало доступным количество органических веществ, сопоставимое с теми, которые производятся из земных источников.[122][123]
Было подсчитано, что поздняя тяжелая бомбардировка могла также эффективно стерилизовать поверхность Земли на глубину до десятков метров. Если бы жизнь эволюционировала глубже, чем это, она также была бы защищена от ранних высоких уровней ультрафиолетового излучения от стадии Т Тельца эволюции Солнца. Моделирование геотермически нагретой океанической коры дает гораздо больше органических веществ, чем те, что были обнаружены в экспериментах Миллера-Юри. В глубокой гидротермальные источники, Эверетт Шок обнаружил, что «существует огромное термодинамическое стремление к образованию органических соединений, поскольку морская вода и гидротермальные жидкости, которые далеки от равновесия, смешиваются и переходят в более стабильное состояние ».[124] Shock обнаружил, что доступная энергия максимальна примерно при 100–150 ° C, именно при температурах, при которых гипертермофильный бактерии и термоацидофильный археи были найдены в основании филогенетическое древо жизни ближе всего к Последний универсальный общий предок (ЛУКА).[125]
Древнейшие свидетельства жизни: палеонтология


Самая ранняя жизнь на Земле существовала более 3,5 Гья (миллиард лет назад),[33][34][35] вовремя Эоархейский Эра, когда после расплавленного Хадейского Эона затвердело достаточное количество корки. Самые ранние вещественные доказательства, обнаруженные на сегодняшний день, состоят из микрофоссилий в Зеленокаменный пояс Нуввуагиттук Северного Квебека, в полосчатая формация железа порождает по крайней мере 3,77 и, возможно, 4,28 Гл.[1][126] Это открытие предполагает, что жизнь возникла очень скоро после образования океанов. Было отмечено, что структура микробов аналогична бактериям, обнаруженным вблизи гидротермальные источники в современную эпоху, и подтвердил гипотезу о том, что абиогенез начался вблизи гидротермальных источников.[41][1]
Биогенный графит был обнаружен в метаосадочных породах 3,7 млрд лет с юго-запада. Гренландия[127] и микробный коврик окаменелости, найденные в песчанике 3,48 Гё из Западная Австралия.[128][129] Свидетельства ранней жизни в скалах из Акилия Остров, недалеко от Пояс Isua супракрустальный на юго-западе Гренландии, датируемые 3,7 Гя, показали биогенные изотопы углерода.[130][131] В других частях супракрустального пояса Исуа включения графита, захваченные внутри гранат кристаллы связаны с другими элементами жизни: кислородом, азотом и, возможно, фосфором в форме фосфат, что дает дополнительные доказательства жизни 3,7 Гя.[132] В Стрелли Пул, в Пилбара региона Западной Австралии убедительные доказательства ранней жизни были обнаружены в пирит -содержащий песчаник на окаменелом пляже, который показал округлые трубчатые клетки, которые окисленный сера фотосинтез при отсутствии кислорода.[133][134][135] Дальнейшие исследования по цирконы из Западной Австралии в 2015 году предположил, что жизнь, вероятно, существовала на Земле по крайней мере 4,1 Гя.[136][137][138]
Концептуальная история до 1960-х годов: биология
Панспермия
Панспермия — это гипотеза который жизнь существует на протяжении всего вселенная, распространяется метеороиды, астероиды, кометы[139] и планетоиды.[140]
Гипотеза панспермии не пытается объяснить, как впервые возникла жизнь, а просто переносит ее происхождение на другую планету или комету. Преимущество внеземного происхождения примитивной жизни состоит в том, что жизнь не обязательно должна формироваться на каждой планете, на которой она встречается, а, скорее, в одном месте, а затем распространяться по всему миру. галактика к другим звездным системам посредством столкновения с кометой и / или метеоритом.[141]Доказательств гипотезы панспермии мало, но они находят некоторую поддержку в исследованиях Марсианские метеориты нашел в Антарктида и в исследованиях экстремофил выживаемость микробов в космических испытаниях.[142][143][144][145]
В августе 2020 года ученые сообщили, что бактерии с Земли, особенно Дейнококк радиодуранс, который очень устойчив к экологические опасности, были обнаружены, что выживали в течение трех лет в космическое пространство, на основе исследований, проведенных на Международная космическая станция.[146][147]
Происхождение жизни установлено сразу после Большого взрыва и распространилось по всей Вселенной.
Крайнее предположение состоит в том, что биохимия жизни могло начаться уже через 17 млн. лет после Большой взрыв, во время обжитая эпоха, и что жизнь может существовать повсюду вселенная.[148][149]
Панспермия, принесенная жизнью с Марса на Землю
Карл Циммер предположил, что химические условия, в том числе присутствие бор, молибден и кислород, необходимый для первоначального производства РНК, возможно, был лучше на раннем Марсе, чем на ранней Земле.[150][151][152] If so, life-suitable molecules originating on Mars may have later migrated to Earth via meteor ejections.
Спонтанное зарождение
General acceptance of spontaneous generation until the 19th century
Traditional religion attributed the origin of life to supernatural deities who created the natural world. Spontaneous generation, the first naturalistic theory of life arising from non-life, goes back to Аристотель и древнегреческая философия, and continued to have support in Western scholarship until the 19th century.[153] Classical notions of spontaneous generation held that certain «lower» or «vermin» animals are generated by decaying organic substances. According to Aristotle, it was readily observable that тля arise from dew on plants, мухи from putrid matter, mice from dirty hay, crocodiles from rotting sunken logs, and so on.[154] A related theory was гетерогенез: that some forms of life could arise from different forms (e.g. bees from flowers).[155] The modern scientist Джон Бернал said that the basic idea of such theories was that life was continuously created as a result of chance events.[156]
In the 17th century, people began to question such assumptions. В 1646 г. Томас Браун опубликовал свой Эпидемическая псевдодоксия (с субтитрами Enquiries into Very many Received Tenets, and commonly Presumed Truths), which was an attack on false beliefs and «vulgar errors.» Его современник, Александр Росс, erroneously refuted him, stating:
To question this [spontaneous generation], is to question Reason, Sense, and Experience: If he doubts of this, let him go to Ægypt, and there he will find the fields swarming with mice begot of the mud of Nylus, to the great calamity of the Inhabitants.[157][158]

Антони ван Левенгук
В 1665 г. Роберт Гук published the first drawings of a микроорганизм. Hooke was followed in 1676 by Антони ван Левенгук, who drew and described microorganisms that are now thought to have been простейшие и бактерии.[159] Many felt the existence of microorganisms was evidence in support of spontaneous generation, since microorganisms seemed too simplistic for половое размножение, и бесполое размножение через деление клеток had not yet been observed. Van Leeuwenhoek took issue with the ideas common at the time that fleas and lice could spontaneously result from гниение, and that frogs could likewise arise from slime. Using a broad range of experiments ranging from sealed and open meat incubation and the close study of insect reproduction he became, by the 1680s, convinced that spontaneous generation was incorrect.[160]
The first experimental evidence against spontaneous generation came in 1668 when Франческо Реди showed that no личинки appeared in meat when flies were prevented from laying eggs. It was gradually shown that, at least in the case of all the higher and readily visible organisms, the previous sentiment regarding spontaneous generation was false. The alternative hypothesis was биогенез: that every living thing came from a pre-existing living thing (omne vivum ex ovo, Latin for «every living thing from an egg»).[161] В 1768 г. Лаззаро Спалланцани продемонстрировал, что микробы were present in the air, and could be killed by boiling. В 1861 г. Луи Пастер performed a series of experiments that demonstrated that organisms such as bacteria and fungi do not spontaneously appear in sterile, nutrient-rich media, but could only appear by invasion from without.
Spontaneous generation considered disproven in the 19th century

Луи Пастер

By the middle of the 19th century, biogenesis had accumulated so much evidence in support that the alternative theory of spontaneous generation had been effectively disproven. Пастер remarked, about a finding of his in 1864 which he considered definitive,
Never will the doctrine of spontaneous generation recover from the mortal blow struck by this simple experiment.[162][163]
gave a mechanism by which life diversified from a few simple organisms to a variety of to complex forms. Today, scientists agree that all current life descends from earlier life, which has become progressively more complex and diverse through Чарльз Дарвин ‘s mechanism of эволюция к естественный отбор.Darwin wrote to Hooker in 1863 stating that,
It is mere rubbish, thinking at present of the origin of life; one might as well think of the origin of matter.
В О происхождении видов, he had referred to life having been «created», by which he «really meant ‘appeared’ by some wholly unknown process», but had soon regretted using the Old Testament term «creation».[нужна цитата ]
Etymology of biogenesis and abiogenesis
Период, термин биогенез is usually credited to either Генри Бастиан или чтобы Томас Хаксли.[164] Bastian used the term around 1869 in an unpublished exchange with Джон Тиндалл to mean «life-origination or commencement». In 1870, Huxley, as new president of the Британская ассоциация развития науки, delivered an address entitled Biogenesis and Abiogenesis.[165] In it he introduced the term биогенез (with an opposite meaning to Bastian’s) as well as абиогенез:
- And thus the hypothesis that living matter always arises by the agency of pre-existing living matter, took definite shape; and had, henceforward, a right to be considered and a claim to be refuted, in each particular case, before the production of living matter in any other way could be admitted by careful reasoners. It will be necessary for me to refer to this hypothesis so frequently, that, to save circumlocution, I shall call it the hypothesis of Биогенез; and I shall term the contrary doctrine—that living matter may be produced by not living matter—the hypothesis of Абиогенез.[165]
Subsequently, in the preface to Bastian’s 1871 book, The Modes of Origin of Lowest Organisms,[166] Bastian referred to the possible confusion with Huxley’s usage and explicitly renounced his own meaning:
- A word of explanation seems necessary with regard to the introduction of the new term Archebiosis. I had originally, in unpublished writings, adopted the word Биогенез to express the same meaning—viz., life-origination or commencement. But in the meantime, the word Биогенез has been made use of, quite independently, by a distinguished biologist [Huxley], who wished to make it bear a totally different meaning. He also introduced the word Абиогенез. I have been informed, however, on the best authority, that neither of these words can—with any regard to the language from which they are derived—be supposed to bear the meanings which have of late been publicly assigned to them. Wishing to avoid all needless confusion, I therefore renounced the use of the word Биогенез, and being, for the reason just given, unable to adopt the other term, I was compelled to introduce a new word, in order to designate the process by which living matter is supposed to come into being, independently of pre-existing living matter.[167]
Since the end of the nineteenth century, ‘evolutive abiogenesis’ means increasing complexity and evolution of matter from inert to living states.[168]
Oparin: Primordial soup hypothesis
There is no single generally accepted model for the origin of life. Scientists have proposed several plausible hypotheses which share some common elements. While differing in details, these hypotheses are based on the framework laid out by Александр Опарин (in 1924) and Джон Холдейн (in 1925), that the first molecules constituting the earliest cells
. . . were synthesized under natural conditions by a slow process of molecular evolution, and these molecules then organized into the first molecular system with properties with biological order».[169]
Oparin and Haldane suggested that the atmosphere of the early Earth may have been chemically reducing in nature, composed primarily of methane (CH4), ammonia (NH3), water (H2O), hydrogen sulfide (H2S), carbon dioxide (CO2) or carbon monoxide (CO), and фосфат (PO43−), with molecular oxygen (O2) и озон (O3) either rare or absent. According to later models, the atmosphere in the late Hadean period consisted largely of nitrogen (N2) and carbon dioxide, with smaller amounts of carbon monoxide, hydrogen (H2), and sulfur compounds;[170] while it did lack molecular oxygen and ozone,[171] it was not as chemically reducing as Oparin and Haldane supposed.
No new notable research or hypothesis on the subject appeared until 1924, when Oparin reasoned that atmospheric oxygen prevents the synthesis of certain organic compounds that are necessary building blocks for life. В его книге Происхождение жизни,[172][173] he proposed (echoing Darwin) that the «spontaneous generation of life» that had been attacked by Pasteur did, in fact, occur once, but was now impossible because the conditions found on the early Earth had changed, and preexisting organisms would immediately consume any spontaneously generated organism. Oparin argued that a «primeval soup» of organic molecules could be created in an oxygenless atmosphere through the action of Солнечный свет. These would combine in ever more complex ways until they formed coacervate капли. These droplets would «расти » by fusion with other droplets, and «воспроизводить » through fission into daughter droplets, and so have a primitive метаболизм in which factors that promote «cell integrity» survive, and those that do not become вымерший. Many modern theories of the origin of life still take Oparin’s ideas as a starting point.
About this time, Haldane suggested that the Earth’s prebiotic oceans (quite different from their modern counterparts) would have formed a «hot dilute soup» in which organic compounds could have formed. Bernal called this idea биопоэз или же биопоэз, the process of living matter evolving from self-replicating but non-living molecules,[156][174] and proposed that biopoiesis passes through a number of intermediate stages.
Роберт Шапиро has summarized the «primordial soup» theory of Oparin and Haldane in its «mature form» as follows:[175]
- The early Earth had a chemically восстановительная атмосфера.
- This atmosphere, exposed to энергия in various forms, produced simple organic compounds («мономеры «).
- These compounds accumulated in a «soup» that may have concentrated at various locations (shorelines, oceanic vents так далее.).
- By further transformation, more complex organic полимеры —and ultimately life—developed in the soup.
Джон Бернал
John Bernal showed that based upon this and subsequent work there is no difficulty in principle in forming most of the molecules we recognize as the necessary molecules for life from their inorganic precursors. The underlying hypothesis held by Oparin, Haldane, Bernal, Miller and Urey, for instance, was that multiple conditions on the primeval Earth favoured chemical reactions that synthesized the same set of complex organic compounds from such simple precursors.Bernal coined the term биопоэз in 1949 to refer to the origin of life.[176] In 1967, he suggested that it occurred in three «stages»:
- the origin of biological monomers
- the origin of biological polymers
- the evolution from molecules to cells
Bernal suggested that evolution commenced between stages 1 and 2. Bernal regarded the third stage, in which biological reactions were incorporated behind a cell’s boundary, as the most difficult. Modern work on the way that клеточные мембраны self-assemble, and the work on micropores in various substrates, may be a key step towards understanding the development of independent free-living cells.[177][178][179]
Эксперимент Миллера – Юри

Стэнли Миллер

Miller–Urey experiment JP
One of the most important pieces of experimental support for the «soup» theory came in 1952. Стэнли Миллер и Гарольд Юри performed an experiment that demonstrated how organic molecules could have spontaneously formed from inorganic precursors under conditions like those posited by the Oparin-Haldane hypothesis. The now-famous Эксперимент Миллера – Юри used a highly reducing mixture of gases—метан, аммиак, и водород, а также водяной пар —to form simple organic monomers such as amino acids.[180] The mixture of gases was cycled through an apparatus that delivered electrical sparks to the mixture. After one week, it was found that about 10% to 15% of the carbon in the system was then in the form of a рацемическая смесь of organic compounds, including amino acids, which are the building blocks of белки. This provided direct experimental support for the second point of the «soup» theory, and it is around the remaining two points of the theory that much of the debate now centers.
A 2011 reanalysis of the saved vials containing the original extracts that resulted from the Miller and Urey experiments, using current and more advanced analytical equipment and technology, has uncovered more biochemicals than originally discovered in the 1950s. One of the more important findings was 23 amino acids, far more than the five originally found.[181]
In November 2020, a team of international scientists reported studies which suggest that the primeval atmosphere of the Earth was much different than the conditions used in the Miller-Urey studies.[182]
Primordial origin of biological molecules: Chemistry
The chemical processes on the pre-biotic early Earth are called chemical evolution. элементы, except for hydrogen and helium, ultimately derive from звездный нуклеосинтез. In 2016, astronomers reported that the very basic chemical ingredients of жизнь — carbon-hydrogen molecule (CH, or метилидиновый радикал ), the carbon-hydrogen positive ion (CH+) and the carbon ion (C+)—are largely the result of ultraviolet light from stars, rather than other forms of radiation from сверхновые и young stars, как думали раньше.[183] Complex molecules, including organic molecules, form naturally both in space and on planets.[24] There are two possible sources of organic molecules on the early Earth:
- Terrestrial origins – organic molecule synthesis driven by impact shocks or by other energy sources (such as UV light, редокс coupling, or electrical discharges; e.g., Miller’s experiments)
- Extraterrestrial origins – formation of organic molecules in interstellar dust clouds, which rain down on planets.[184][185] (Видеть псевдопанспермия )
An organic compound is any member of a large class of gaseous, liquid, or solid chemicals whose molecules contain carbon. Carbon is the fourth most abundant element in the Universe by mass after hydrogen, гелий, and oxygen.[186] Carbon is abundant in the Sun, stars, comets, and in the атмосферы of most planets.[187] Organic compounds are relatively common in space, formed by «factories of complex molecular synthesis» which occur in молекулярные облака и circumstellar envelopes, and chemically evolve after reactions are initiated mostly by ионизирующего излучения.[24][188][189][190] На основе computer model studies, the complex organic molecules necessary for life may have formed on dust grains in the protoplanetary disk surrounding the Sun before the formation of the Earth.[191] According to the computer studies, this same process may also occur around other stars that acquire planets.[191]
Аминокислоты
NASA announced in 2009 that scientists had identified another fundamental chemical building block of life in a comet for the first time, glycine, an amino acid, which was detected in material ejected from comet Wild 2 in 2004 and grabbed by NASA’s Звездная пыль зонд. Glycine has been detected in meteorites before. Carl Pilcher, who leads the Институт астробиологии НАСА прокомментировал это
The discovery of glycine in a comet supports the idea that the fundamental building blocks of life are prevalent in space, and strengthens the argument that life in the universe may be common rather than rare.[192]
Comets are encrusted with outer layers of dark material, thought to be a деготь -like substance composed of complex organic material formed from simple carbon compounds after reactions initiated mostly by ionizing radiation. It is possible that a rain of material from comets could have brought significant quantities of such complex organic molecules to Earth.[193][194][195] Amino acids which were formed extraterrestrially may also have arrived on Earth via comets.[49] It is estimated that during the Late Heavy Bombardment, meteorites may have delivered up to five million тонны of organic prebiotic elements to Earth per year.[49]
Гипотеза мира ПАУ
Полициклические ароматические углеводороды (PAH) are the most common and abundant of the known polyatomic molecules in the наблюдаемая вселенная, and are considered a likely constituent of the primordial sea.[196][197][198] In 2010, PAHs, have been detected in туманности.[199]

Polycyclic aromatic hydrocarbons (PAH) are known to be abundant in the universe,[196][197][198] в том числе в межзвездная среда, in comets, and in meteorites, and are some of the most complex molecules so far found in space.[187]
Other sources of complex molecules have been postulated, including extraterrestrial stellar or interstellar origin. For example, from spectral analyses, organic molecules are known to be present in comets and meteorites. In 2004, a team detected traces of PAHs in a nebula.[200] In 2010, another team also detected PAHs, along with fullerenes, in nebulae.[199] The use of PAHs has also been proposed as a precursor to the RNA world in the PAH world hypothesis.[201] В Космический телескоп Спитцера has detected a star, HH 46-IR, which is forming by a process similar to that by which the Sun formed. In the disk of material surrounding the star, there is a very large range of molecules, including cyanide compounds, углеводороды, and carbon monoxide. In 2012, NASA scientists reported that PAHs, subjected to interstellar medium conditions, are transformed, through hydrogenation, оксигенация и гидроксилирование, to more complex organics—»a step along the path toward amino acids and nucleotides, the raw materials of proteins and DNA, respectively.»[202][203] Далее, в результате этих превращений ПАУ теряют свою спектроскопическая подпись что могло быть одной из причин «отсутствия обнаружения ПАУ в межзвездный лед grains, particularly the outer regions of cold, dense clouds or the upper molecular layers of protoplanetary disks.»[202][203]
NASA maintains a database for tracking PAHs in the universe.[187][204] More than 20% of the carbon in the universe may be associated with PAHs,[187] possible starting materials for the formation of life. PAHs seem to have been formed shortly after the Big Bang, are widespread throughout the universe,[196][197][198] и связаны с новые звезды и экзопланеты.[187]
Нуклеооснования
Observations suggest that the majority of organic compounds introduced on Earth by interstellar dust particles are considered principal agents in the formation of complex molecules, thanks to their peculiar surface-catalytic виды деятельности.[205][206] Studies reported in 2008, based on 12C /13C isotopic ratios of organic compounds found in the Murchison meteorite, suggested that the RNA component uracil and related molecules, including ксантин, were formed extraterrestrially.[207][208] In 2011, a report based on НАСА studies of meteorites found on Earth was published suggesting DNA components (adenine, guanine and related organic molecules) were made in outer space.[205][209][210] Scientists also found that the космическая пыль permeating the universe contains complex organics («amorphous organic solids with a mixed ароматный –алифатический structure») that could be created naturally, and rapidly, by stars.[211][212][213] Сун Квок из Университет Гонконга suggested that these compounds may have been related to the development of life on Earth said that «If this is the case, life on Earth may have had an easier time getting started as these organics can serve as basic ingredients for life.»[211]
The sugar glycolaldehyde

Glycolaldehyde, the first example of an interstellar sugar molecule, was detected in the star-forming region near the centre of our galaxy. It was discovered in 2000 by Jes Jørgensen and Jan Hollis.[214] In 2012, Jørgensen’s team reported the detection of glycolaldehyde in a distant star system. Молекула была обнаружена вокруг протозвездный двоичный IRAS 16293-2422 400 световых лет с Земли.[215][216][217] Glycolaldehyde is needed to form RNA, which is similar in function to DNA. These findings suggest that complex organic molecules may form in stellar systems prior to the formation of planets, eventually arriving on young planets early in their formation.[218][219] Because sugars are associated with both metabolism and the генетический код, two of the most basic aspects of life, it is thought the discovery of extraterrestrial sugar increases the likelihood that life may exist elsewhere in our galaxy.[214]
Polyphosphates
A problem in most scenarios of abiogenesis is that the thermodynamic equilibrium of amino acid versus peptides is in the direction of separate amino acids. What has been missing is some force that drives polymerization. The resolution of this problem may well be in the properties of полифосфаты.[220][221] Polyphosphates are formed by polymerization of ordinary monophosphate ions PO43-. Several mechanisms of organic molecule synthesis have been investigated. Polyphosphates cause polymerization of amino acids into peptides. They are also logical precursors in the synthesis of such key biochemical compounds as аденозинтрифосфат (АТФ). A key issue seems to be that calcium reacts with soluble phosphate to form insoluble фосфат кальция (апатит ), so some plausible mechanism must be found to keep calcium ions from causing precipitation of phosphate. There has been much work on this topic over the years, but an interesting new idea is that meteorites may have introduced reactive phosphorus species on the early Earth.[222]Based on recent computer model studies, то сложные органические молекулы necessary for life may have formed in the протопланетный диск из пылинки окружающий солнце before the formation of the Earth.[191][223] According to the computer studies, this same process may also occur around other звезды that acquire планеты. (Также см Extraterrestrial organic molecules ).
The accumulation and concentration of organic molecules on a planetary surface is also considered an essential early step for the origin of life.[43] Identifying and understanding the mechanisms that led to the production of prebioticmolecules in various environments is critical for establishing the inventory of ingredients from which life originated on Earth, assuming that the abiotic production of molecules ultimately influenced the selection of molecules from which life emerged.[43]
In 2019, scientists reported detecting, for the first time, молекулы сахара, включая рибоза, в meteorites, предполагая, что химические процессы на астероиды может производить некоторые принципиально важные био-ингредиенты, важные для жизнь, и поддерживая понятие Мир РНК prior to a DNA-based origin of life on Earth, and possibly, as well, the notion of панспермия.[224][219]
Chemical synthesis in the laboratory
As early as the 1860s, experiments have demonstrated that biologically relevant molecules can be produced from interaction of simple carbon sources with abundant inorganic catalysts.
Fox proteinoids
In trying to uncover the intermediate stages of abiogenesis mentioned by Bernal, Сидни Фокс in the 1950s and 1960s studied the spontaneous formation of пептид structures (small chains of amino acids) under conditions that might plausibly have existed early in Earth’s history. In one of his experiments, he allowed amino acids to dry out as if puddled in a warm, dry spot in prebiotic conditions: In an experiment to set suitable conditions for life to form, Fox collected volcanic material from a шлаковый конус в Гавайи. He discovered that the temperature was over 100 C just 4 inches (100 mm) beneath the surface of the cinder cone, and suggested that this might have been the environment in which life was created—molecules could have formed and then been washed through the loose volcanic ash into the sea. He placed lumps of lava over amino acids derived from methane, ammonia and water, sterilized all materials, and baked the lava over the amino acids for a few hours in a glass oven. A brown, sticky substance formed over the surface, and when the lava was drenched in sterilized water, a thick, brown liquid leached out. He found that, as they dried, the amino acids formed long, often cross-linked, thread-like, submicroscopic полипептид молекулы.[225]
Сахара
In particular, experiments by Бутлеров (в formose reaction ) showed that tetroses, pentoses, and hexoses are produced when formaldehyde is heated under basic conditions with divalent metal ions like calcium. The reaction was scrutinized and subsequently proposed to be autocatalytic by Breslow in 1959.
Нуклеооснования
Similar experiments (see below) demonstrate that nucleobases like guanine and adenine could be synthesized from simple carbon and nitrogen sources like hydrogen cyanide and ammonia.
Формамид produces all four ribonucleotides and other biological molecules when warmed in the presence of various terrestrial minerals. Formamide is ubiquitous in the Universe, produced by the reaction of water and цианистый водород (HCN). It has several advantages as a biotic precursor, including the ability to easily become concentrated through the evaporation of water.[226][227] Although HCN is poisonous, it only affects аэробные организмы (эукариоты and aerobic bacteria), which did not yet exist. It can play roles in other chemical processes as well, such as the synthesis of the amino acid глицин.[49]
In March 2015, NASA scientists reported that, for the first time, complex DNA and RNA organic compounds of life, including uracil, cytosine and тимин, have been formed in the laboratory under outer space conditions, using starting chemicals, such as pyrimidine, found in meteorites. Pyrimidine, like PAHs, the most carbon-rich chemical found in the Universe, may have been formed in красный гигант stars or in interstellar dust and gas clouds.[228] A group of Czech scientists reported that all four RNA-bases may be synthesized from formamide in the course of high-energy density events like extraterrestrial impacts.[229]
Use of high temperature
In 1961, it was shown that the nucleic acid пурин основание аденин can be formed by heating aqueous ammonium cyanide решения.[230]
Use of low (freezing) temperature
Other pathways for synthesizing bases from inorganic materials were also reported.[231] Orgel and colleagues have shown that freezing temperatures are advantageous for the synthesis of purines, due to the concentrating effect for key precursors such as hydrogen cyanide.[232] Research by Miller and colleagues suggested that while adenine and гуанин require freezing conditions for synthesis, цитозин и урацил may require boiling temperatures.[233] Research by the Miller group notes the formation of seven different amino acids and 11 types of азотистые основания in ice when ammonia and цианид were left in a freezer from 1972 to 1997.[234][235] Other work demonstrated the formation of s-triazines (alternative nucleobases), пиримидины (including cytosine and uracil), and adenine from urea solutions subjected to freeze-thaw cycles under a reductive atmosphere (with spark discharges as an energy source).[236] The explanation given for the unusual speed of these reactions at such a low temperature is eutectic freezing. As an ice crystal forms, it stays pure: only molecules of water join the growing crystal, while impurities like salt or cyanide are excluded. These impurities become crowded in microscopic pockets of liquid within the ice, and this crowding causes the molecules to collide more often. Mechanistic exploration using quantum chemical methods provide a more detailed understanding of some of the chemical processes involved in chemical evolution, and a partial answer to the fundamental question of molecular biogenesis.[237]
Use of less-reducing gas in Miller–Urey experiment
At the time of the Miller–Urey experiment, scientific consensus was that the early Earth had a reducing atmosphere with compounds relatively rich in hydrogen and poor in oxygen (e.g., CH4 и NH3 as opposed to CO2 и диоксид азота (НЕТ2)). However, current scientific consensus describes the primitive atmosphere as either weakly reducing or neutral[238][239] (смотрите также Oxygen Catastrophe ). Such an atmosphere would diminish both the amount and variety of amino acids that could be produced, although studies that include утюг и карбонат minerals (thought present in early oceans) in the experimental conditions have again produced a diverse array of amino acids.[238] Other scientific research has focused on two other potential reducing environments: космическое пространство and deep-sea thermal vents.[240][241][242]
Synthesis based on hydrogen cyanide
A research project completed in 2015 by Джон Сазерленд and others found that a network of reactions beginning with hydrogen cyanide and hydrogen sulfide, in streams of water irradiated by UV light, could produce the chemical components of proteins and lipids, as well as those of RNA,[243][244] while not producing a wide range of other compounds.[245] The researchers used the term «cyanosulfidic» to describe this network of reactions.[244]
Issues during laboratory synthesis
The spontaneous formation of complex polymers from abiotically generated monomers under the conditions posited by the «soup» theory is not at all a straightforward process. Besides the necessary basic organic monomers, compounds that would have prohibited the formation of polymers were also formed in high concentration during the Miller–Urey and Хоан Оро эксперименты.[246] The Miller–Urey experiment, for example, produces many substances that would react with the amino acids or terminate their coupling into peptide chains.[247]
Автокатализ
Autocatalysts are substances that catalyze the production of themselves and therefore are «molecular replicators.» The simplest self-replicating chemical systems are autocatalytic, and typically contain three components: a product molecule and two precursor molecules. The product molecule joins together the precursor molecules, which in turn produce more product molecules from more precursor molecules. The product molecule catalyzes the reaction by providing a complementary template that binds to the precursors, thus bringing them together. Such systems have been demonstrated both in biological макромолекулы and in small organic molecules.[248][249] Systems that do not proceed by template mechanisms, such as the self-reproduction of мицеллы и пузырьки, have also been observed.[249]
It has been proposed that life initially arose as autocatalytic chemical networks.[250] Британский этолог Ричард Докинз wrote about autocatalysis as a potential explanation for the origin of life in his 2004 book Рассказ предков.[251] In his book, Dawkins cites experiments performed by Юлиус Ребек and his colleagues in which they combined amino adenosine and pentafluorophenyl esters with the autocatalyst amino adenosine triacid ester (AATE). One product was a variant of AATE, which catalyzed the synthesis of themselves. This experiment demonstrated the possibility that autocatalysts could exhibit competition within a population of entities with heredity, which could be interpreted as a rudimentary form of natural selection.[252][253]
Encapsulation: morphology
Encapsulation without a membrane
Oparin’s coacervate
Membraneless polyester droplets
Researchers Tony Jia and Kuhan Chandru[254] have proposed that membraneless polyesters droplets could have been significant in the Origins of Life.[255] Given the «messy» nature of prebiotic chemistry,[256][257] the spontaneous generation of these combinatorial droplets may have played a role in early cellularization before the innovation of lipid vesicles. Protein function within and RNA function in the presence of certain polyester droplets was shown to be preserved within the droplets. Additionally, the droplets have scaffolding ability, by allowing lipids to assemble around them that may have prevented leakage of genetic materials.
Proteinoid microspheres
Fox observed in the 1960s that the proteinoids that he had synthesized could form cell-like structures that have been named «proteinoid microspheres «.[225]
The amino acids had combined to form proteinoids, and the proteinoids had combined to form small globules that Fox called «microspheres». His proteinoids were not cells, although they formed clumps and chains reminiscent of цианобактерии, but they contained no functional нуклеиновые кислоты or any encoded information. Based upon such experiments, Колин Питтендрай stated in 1967 that «laboratories will be creating a living cell within ten years,» a remark that reflected the typical contemporary naivety about the complexity of cell structures.[258]
Lipid world
В lipid world theory postulates that the first self-replicating object was липид -подобно.[259][260] It is known that phospholipids form липидные бислои in water while under agitation—the same structure as in cell membranes. These molecules were not present on early Earth, but other амфифильный long-chain molecules also form membranes. Кроме того, эти тела могут расширяться (за счет введения дополнительных липидов), а при чрезмерном расширении могут подвергаться спонтанному расщеплению, которое сохраняет одинаковый размер и состав липидов в двух потомство. Основная идея этой теории состоит в том, что молекулярный состав липидных тел является предварительным способом хранения информации, а эволюция привела к появлению полимерных структур, таких как РНК или ДНК, которые могут благоприятно хранить информацию. Исследования пузырьков из потенциально пребиотических амфифилов до сих пор ограничивались системами, содержащими один или два типа амфифилов. Это в отличие от результатов моделирования пребиотических химических реакций, которые обычно приводят к очень гетерогенным смесям соединений.[261]В рамках гипотезы о липидной бислойной мембране, состоящей из смеси различных различных амфифильных соединений, существует возможность огромного числа теоретически возможных комбинаций в расположении этих амфифилов в мембране. Среди всех этих возможных комбинаций конкретное локальное расположение мембраны способствовало бы образованию гиперцикла,[262][263] на самом деле положительный Обратная связь состоит из двух общих катализаторов, представленных участком мембраны и определенным соединением, заключенным в везикулу. Такие пары сайт / соединение передаются дочерним везикулам, что приводит к появлению отдельных родословная пузырьков, которые позволили бы дарвиновскому естественному отбору.[264]
Протоклетки
Протоклетка — это самоорганизованная, самоупорядоченная сферическая совокупность липиды предлагается как ступенька к зарождению жизни.[261] Центральный вопрос эволюции заключается в том, как простые протоклетки впервые возникли и различались по репродуктивному вкладу в следующее поколение, управляющее эволюцией жизни. Хотя функциональная протоклетка еще не была достигнута в лабораторных условиях, есть ученые, которые думают, что цель вполне достижима.[265][266][267]
Самостоятельная сборка пузырьки являются важными компонентами примитивных клеток.[261] В второй закон термодинамики требует, чтобы Вселенная двигалась в направлении, в котором энтропия возрастает, но жизнь отличается большой степенью организации. Следовательно, необходима граница для разделения жизненные процессы из неживой материи.[268] Исследователи Ирен Чен и Шостак, среди прочих, предполагают, что простые физико-химические свойства элементарных протоклеток могут привести к существенному клеточному поведению, включая примитивные формы дифференциальной конкуренции воспроизводства и накопления энергии. Такие совместные взаимодействия между мембраной и ее инкапсулированным содержимым могут значительно упростить переход от простых реплицирующихся молекул к истинным клеткам.[266] Кроме того, конкуренция за мембранные молекулы будет благоприятствовать стабилизированным мембранам, что предполагает селективное преимущество для эволюции сшитых жирных кислот и даже фосфолипиды сегодня.[266] Такой микрокапсулирование позволит метаболизм внутри мембраны, обмен маленькими молекулами, но предотвращение прохождения через нее крупных веществ.[269] К основным преимуществам инкапсуляции можно отнести повышенную растворимость содержащегося в капсуле груза и накопителя энергии в виде электрохимический градиент.
Другой подход к понятию протоклетки касается термина «хемотон «(сокращение от ‘химический автомат ‘), который относится к абстрактной модели фундаментальной единицы жизни, представленной Венгерский биолог-теоретик Тибор Ганти.[270] Это старейшая из известных вычислительных аннотаций протоклетки. Ганти задумал основную идею в 1952 году и сформулировал ее в 1971 году в своей книге. Принципы жизни (Первоначально написано на венгерском языке и переведено на английский только в 2003 году). Он предположил, что хемотон является изначальным предком всех организмов, или последний универсальный общий предок.[271]
Основное предположение хемотонной модели состоит в том, что жизнь должна иметь три основных свойства: метаболизм, самовоспроизведение, а билипидная мембрана.[272] Функции метаболизма и репликации вместе образуют автокаталитический подсистема необходима для основных функций жизни, и мембрана включает эту подсистему, чтобы отделить ее от окружающей среды. Следовательно, любая система, обладающая такими свойствами, может считаться живой и подвергаться естественный отбор и содержат самоподдерживающуюся клеточную информацию. Некоторые считают эту модель значительным вкладом в зарождение жизни, поскольку она обеспечивает философию эволюционные единицы.[273]
Тем не менее, исследование 2012 г., проведенное Мулкиджаняном из Оснабрюкский университет, предполагает, что внутренние бассейны с конденсированным и охлажденным геотермальным паром обладают идеальными характеристиками для происхождения жизни.[274] Ученые подтвердили в 2002 году, что добавлением монтмориллонит глины в раствор мицелл жирных кислот (липидных сфер), глина ускоряла скорость образования пузырьков в 100 раз.[267] Более того, недавние исследования показали, что повторяющиеся действия дегидратации и регидратации захватывают биомолекулы, такие как РНК, внутри липидных протоклеток, обнаруженных в горячих источниках, и создают необходимые предпосылки для эволюции путем естественного отбора.[275]
Образование липидных пузырьков в пресной воде
Брюс Дамер и Дэвид Димер пришли к выводу, что клеточные мембраны не может образоваться в соленом морская вода, и, следовательно, должны были происходить из пресной воды. До образования континентов единственной сушей на Земле были вулканические острова, где дождевая вода образовывала пруды, где липиды могли образовывать первые стадии на пути к клеточным мембранам. Предполагается, что эти предшественники настоящих ячеек вели себя скорее как суперорганизм а не отдельные структуры, где пористые мембраны будут содержать молекулы, которые просачиваются наружу и попадают в другие протоклетки. Только когда настоящие клетки эволюционируют, они постепенно адаптируются к более соленой среде и выходят в океан.[276]
Везикулы, состоящие из смесей РНК-подобных биохимических веществ.
Другая модель протоклетки — это Jeewanu. Впервые синтезированный в 1963 году из простых минералов и основных органических веществ под воздействием солнечного света, он до сих пор обладает некоторыми метаболическими способностями, а именно: полупроницаемая мембрана, аминокислоты, фосфолипиды, углеводы и РНК-подобные молекулы.[277][278] Однако природа и свойства Дживану еще предстоит выяснить.
Электростатические взаимодействия, индуцированные короткими, положительно заряженными, гидрофобными пептидами, содержащими 7 аминокислот в длину или меньше, могут прикреплять РНК к мембране везикул, основной клеточной мембране.[279][280]
Осадки сульфидов металлов
Уильям Мартин и Майкл Рассел предложили
. . . . что жизнь развивалась в структурированных осадках моносульфида железа в гидротермальном насыпи фильтрующего участка при окислительно-восстановительном потенциале, pH и температурном градиенте между гидротермальным флюидом, богатым сульфидами, и содержащими железо (II) водами на дне океана Хадей. Естественно возникающая трехмерная компартментация, наблюдаемая в окаменелых осадках сульфидов металлов в местах утечки, указывает на то, что эти неорганические компартменты были предшественниками клеточных стенок и мембран, обнаруженных у свободноживущих прокариот. Известная способность FeS и NiS катализировать синтез ацетил-метилсульфида из монооксида углерода и метилсульфида, составляющих гидротермальный флюид, указывает на то, что предбиотические синтезы происходили на внутренних поверхностях этих секций со стенками из сульфидов металлов, … «[281]
Соответствующие геологические среды
Маленький пруд Дарвина
Ранняя концепция, что жизнь возникла из неживой материи на медленных стадиях, появилась в Герберт Спенсер книга 1864–1867 гг. Принципы биологии. В 1879 г. Уильям Тернер Тизелтон-Дайер говорится об этом в статье «О самозарождении и эволюции». 1 февраля 1871 г. Чарльз Дарвин написал об этих публикациях Джозеф Хукер, и изложил свои предположения,[282][283][284] предполагая, что первоначальная искра жизни могла начаться в
теплый маленький пруд со всеми видами аммиака и фосфорных солей, светом, теплом, электричеством и т. д. протеин соединение было химически образовано, готовым претерпеть еще более сложные изменения.
Он продолжил объяснять, что
в настоящее время такая материя будет немедленно поглощена или поглощена, чего не было бы до образования живых существ.
Дарвин 1887, п.18:
Часто говорят, что сейчас существуют все условия для первого появления живого организма, которые могли когда-либо существовать. Но если бы (и о! Какое большое если!) Мы могли бы зачать ребенка в каком-нибудь теплом маленьком пруду со всеми видами аммиака и фосфорных солей, светом, теплом, электричеством и т. Д. протеин соединение было химически сформировано, готовым подвергнуться еще более сложным изменениям, в настоящее время такое вещество будет немедленно поглощено или поглощено, чего не было бы до образования живых существ.
— Дарвин, 1 февраля 1871 г.
Более поздние исследования, проведенные в 2017 году, подтверждают мнение о том, что жизнь могла возникнуть сразу после того, как Земля была сформирована в виде молекул РНК, выходящих из «теплых маленьких водоемов».[285]
Вулканические горячие источники и гидротермальные источники, мелкие или глубокие
Мартин Бразье показал, что ранние окаменелости произошли из горячего мира газов, таких как метан, аммиак, углекислый газ и сероводород, которые токсичны для многих из нас.[286] Другой анализ обычного тройного древа жизни показывает теплолюбивость и гипертермофильность. бактерии и археи находятся ближе всего к корню, что позволяет предположить, что жизнь могла развиваться в жаркой среде.[287]
Глубоководные гидротермальные источники

Глубоководный или щелочной гидротермальный источник, теория утверждает, что жизнь могла начаться в подводных гидротермальных источниках,[288][289] Мартин и Рассел предложили
что жизнь развивалась в структурированных осадках моносульфида железа в гидротермальном насыпи фильтрующего участка при окислительно-восстановительном потенциале, pH и температурном градиенте между гидротермальным флюидом, богатым сульфидами, и содержащими железо (II) водами на дне океана Хадей. Естественно возникающая трехмерная компартментация, наблюдаемая в окаменелых осадках сульфидов металлов в местах утечки, указывает на то, что эти неорганические компартменты были предшественниками клеточных стенок и мембран, обнаруженных у свободноживущих прокариот. Известная способность FeS и NiS катализировать синтез ацетил-метилсульфида из монооксида углерода и метилсульфида, составляющих гидротермальный флюид, указывает на то, что предбиотические синтезы происходили на внутренних поверхностях этих секций со стенками из сульфидов металлов, …[281]
Они образуются там, где богатые водородом жидкости выходят из-под морского дна в результате серпентинизация ультра-мафический оливин с морской водой и границей pH с океанской водой, богатой углекислым газом. Отверстия образуют устойчивый источник химической энергии, полученный в результате окислительно-восстановительных реакций, в которых доноры электронов (молекулярный водород) реагируют с акцепторами электронов (диоксид углерода); видеть Теория мира железо-сера. Это очень экзотермические реакции.[288][c]
Рассел продемонстрировал, что щелочные вентили создают абиогенный движущая сила протона (PMF) хемиосмотический градиент[281] в которых условия идеальны для абиогенного инкубатория на всю жизнь. Их микроскопические отсеки «обеспечивают естественные средства для концентрации органических молекул», состоящих из железо-серных минералов, таких как макинавит, наделил эти минеральные клетки каталитическими свойствами, предусмотренными Гюнтер Вехтерсхойзер.[290] Это движение ионов через мембрану зависит от комбинации двух факторов:
- Распространение сила, вызванная градиентом концентрации — все частицы, включая ионы, имеют тенденцию диффундировать от более высокой концентрации к более низкой.
- Электростатическая сила, вызванная градиентом электрического потенциала -катионы подобно протоны ЧАС+ имеют тенденцию рассеивать электрический потенциал, анионы в обратном направлении.
Эти два градиента, вместе взятые, можно выразить как электрохимический градиент, обеспечивая энергию для абиогенного синтеза. Движущая сила протона может быть описана как мера потенциальной энергии, запасенной как комбинация протонов и градиентов напряжения на мембране (различия в концентрации протонов и электрического потенциала).
Шостак предположил, что геотермальная активность предоставляет большие возможности для зарождения жизни в открытых озерах, где происходит накопление полезных ископаемых. В 2010 году на основе спектрального анализа морской и горячей минеральной воды Игнат Игнатов и Олег Мосин продемонстрировали, что жизнь, возможно, возникла преимущественно в горячей минеральной воде. Горячая минеральная вода, содержащая бикарбонат и кальций Ионы имеет наиболее оптимальный диапазон.[291] Этот случай похож на зарождение жизни в гидротермальных источниках, но с ионами бикарбоната и кальция в горячей воде. Эта вода имеет pH 9–11, и в морской воде возможны реакции. В соответствии с Мелвин Кэлвин, определенные реакции конденсации-дегидратации аминокислот и нуклеотидов в отдельных блоках пептидов и нуклеиновых кислот могут происходить в первичной гидросфере с pH 9–11 на более позднем этапе эволюции.[292] Некоторые из этих соединений, например синильная кислота (HCN) были доказаны в экспериментах Миллера. Это среда, в которой строматолиты были созданы. Дэвид Уорд из Государственный университет Монтаны описали образование строматолитов в горячей минеральной воде на Йеллоустонский Национальный Парк. Строматолиты выживают в горячей минеральной воде и вблизи районов с вулканической активностью.[293] В море у гейзеров горячей минеральной воды развивались процессы. В 2011 году Тадаси Сугавара из Токийский университет создал протоклетку в горячей воде.[294]
Экспериментальные исследования и компьютерное моделирование показывают, что поверхности минеральных частиц внутри гидротермальных источников обладают каталитическими свойствами, аналогичными свойствам ферментов, и способны создавать простые органические молекулы, такие как метанол (CH3ОН) и муравьиной, уксусный и пировиноградный кислота из растворенного CO2 в воде.[295][296]
Исследование, опубликованное выше Мартином в 2016 году, подтверждает тезис о том, что жизнь возникла в гидротермальных источниках,[297][298] что спонтанная химия в земной коре, вызванная взаимодействиями породы и воды при неравновесном термодинамическом уровне, лежала в основе происхождения жизни[299][300] и что основоположники архей и бактерий были H2-зависимыми автотрофами, которые использовали CO2 в качестве конечного акцептора в энергетическом метаболизме.[301] Мартин предполагает, основываясь на этих доказательствах, что LUCA «возможно, выживание сильно зависело от геотермальной энергии вентиляционного канала».[302]
Колеблющиеся гидротермальные бассейны на вулканических островах или протоконтинентах
Мулкиджанян и соавторы считают, что морская среда не обеспечивала ионный баланс и состав, универсальные для клеток, а также ионный баланс, необходимый для основных белков и рибозимов, обнаруженных практически во всех живых организмах, особенно в отношении K+/ Na+ отношение, Mn2+, Zn2+ и концентрации фосфатов. Единственная известная среда, которая имитирует необходимые условия на Земле, находится в земных гидротермальных бассейнах, питаемых паровыми отверстиями.[288] Кроме того, минеральные отложения в этих средах в бескислородной атмосфере будут иметь подходящий pH (в отличие от текущих бассейнов в насыщенной кислородом атмосфере), будут содержать осадки сульфидных минералов, которые блокируют вредное УФ-излучение, иметь циклы смачивания / сушки, которые концентрируют растворы субстрата до приемлемых концентраций. к самопроизвольному образованию полимеров нуклеиновых кислот, полиэфиров[303] и депсипептиды,[304] как химическими реакциями в гидротермальной среде, так и воздействием УФ-излучение при транспортировке из вентиляционных отверстий в соседние бассейны. Их гипотетические предбиотические среды аналогичны наиболее часто предполагаемым глубоководным жерловым средам, но добавляют дополнительные компоненты, которые помогают объяснить особенности, обнаруженные при реконструкциях Последний универсальный общий предок (LUCA) всех живых организмов.[305]
Колин-Гарсия и другие. (2016) обсуждают преимущества и недостатки гидротермальных источников как примитивных сред.[288] Они упоминают, что экзэргонические реакции в таких системах могли быть источником свободной энергии, которая способствовала химическим реакциям, в дополнение к их высокому минералогическому разнообразию, которое подразумевает индукцию важных химических градиентов, что способствует взаимодействию между донорами и акцепторами электронов. Колин-Гарсия и другие. (2016) также обобщают ряд экспериментов, предложенных для проверки роли гидротермальных источников в синтезе пребиотиков.[288]
Вулканический пепел в океане
Джеффри В. Хоффманн утверждал, что сложное событие нуклеации как источник жизни, включающее как полипептиды, так и нуклеиновую кислоту, совместимо со временем и пространством, доступными в примитивных океанах Земли.[306] Хоффманн предполагает, что вулканический пепел может иметь множество случайных форм, необходимых в постулируемом событии комплексного зарождения. Этот аспект теории можно проверить экспериментально.
Раскаленная биосфера золота
В 1970-е годы Томас Голд предложил теорию о том, что жизнь сначала зародилась не на поверхности Земли, а на несколько километров ниже поверхности. Утверждается, что открытие микробной жизни под поверхностью другого тела в нашей Солнечной системе сделало бы эту теорию достоверной. Голд также утверждал, что для выживания необходима тонкая струйка пищи из глубокого, недоступного источника, потому что жизнь, возникающая в луже органического материала, вероятно, поглотит всю свою пищу и вымрет. Теория Голда состоит в том, что поток такой пищи происходит из-за выделения газа первичного метана из мантии Земли; Более традиционное объяснение питания глубоких микробов (вдали от углеродных соединений осадка) состоит в том, что организмы питаться водородом высвобождается при взаимодействии воды и (восстановленных) соединений железа в горных породах.
Гипотеза радиоактивного пляжа
Захари Адам утверждает, что приливные процессы, происходившие в то время, когда Луна была намного ближе, могли иметь концентрированные частицы уран и другие радиоактивные элементы на отметке высокого уровня воды на исконных пляжах, где они, возможно, были ответственны за создание строительных блоков жизни.[307] Согласно компьютерным моделям,[308] месторождение таких радиоактивных материалов может показать то же самое самоподдерживающаяся ядерная реакция как это найдено в Окло пласт урановой руды в Габон. Такой радиоактивный пляжный песок мог обеспечивать достаточно энергии для образования органических молекул, таких как аминокислоты и сахара, из ацетонитрил в воде. Радиоактивный монацит Материал также высвободил растворимый фосфат в области между песчинками, что сделало его биологически «доступным». Таким образом, по мнению Адама, аминокислоты, сахара и растворимые фосфаты могли производиться одновременно. Радиоактивный актиниды, оставленный реакцией в некоторой концентрации, мог быть частью металлоорганические комплексы. Эти комплексы могли быть важными ранними катализаторами жизненных процессов.
Джон Парнелл предположил, что такой процесс мог бы стать частью «тигля жизни» на ранних стадиях любой ранней влажной скалистой планеты, пока планета достаточно велика, чтобы сформировать систему тектоники плит, которая приносит радиоактивные минералы. поверхность. Считается, что на ранней Земле было много меньших плит, поэтому она могла обеспечить подходящую среду для таких процессов.[309]
Происхождение метаболизма: физиология
Различные формы жизни с процессами переменного происхождения могли появиться квазиодновременно в раннем история Земли.[310] Другие формы могут быть вымершими (оставив отличительные окаменелости благодаря разной биохимии — например, гипотетические типы биохимии ). Было предложено:
Первыми организмами были самовоспроизводящиеся глины, богатые железом, которые связывали углекислый газ в щавелевую и другие дикарбоновые кислоты. Эта система реплицирующихся глин и их метаболический фенотип затем эволюционировали в богатую сульфидами область горячего источника, приобретая способность фиксировать азот. Наконец, фосфат был включен в развивающуюся систему, что позволило синтезировать нуклеотиды и фосфолипиды. Если биосинтез повторяет биопоэз, то синтез аминокислот предшествовал синтезу пуриновых и пиримидиновых оснований. Кроме того, полимеризация сложных тиоэфиров аминокислот в полипептиды предшествовала направленной полимеризации сложных эфиров аминокислот полинуклеотидами.[311]
Реакции, подобные метаболизму, могли произойти естественным образом в ранних океанах, до того, как появились первые организмы.[20][312] Метаболизм может предшествовать возникновению жизни, которая могла возникнуть в результате химических условий в самых ранних океанах. Реконструкции в лабораториях показывают, что некоторые из этих реакций могут производить РНК, а некоторые другие напоминают два основных реакционных каскада метаболизма: гликолиз и пентозофосфатный путь, которые предоставляют необходимые предшественники нуклеиновых кислот, аминокислот и липидов.[312]
Гипотеза глины
Монтмориллонит, обильный глина, является катализатором полимеризации РНК и образования мембран из липидов.[313] Модель происхождения жизни с использованием глины была предложена Александром Кэрнс-Смитом в 1985 году и исследована в качестве вероятного механизма несколькими учеными.[314] Гипотеза глины постулирует, что сложные органические молекулы постепенно возникали на ранее существовавших, неорганических поверхностях репликации кристаллов силиката в растворе.
На Политехнический институт Ренсселера Исследования Джеймса Ферриса также подтвердили, что минералы монтмориллонитовой глины катализируют образование РНК в водном растворе, соединяя нуклеотиды с образованием более длинных цепей.[315]
В 2007 году Барт Кар из Вашингтонский университет и коллеги сообщили о своих экспериментах, в которых проверялась идея о том, что кристаллы могут действовать как источник передаваемой информации, используя кристаллы гидрофталат калия. «Материнские» кристаллы с дефектами раскалывались и использовались в качестве затравки для выращивания «дочерних» кристаллов из раствора. Затем они исследовали распределение дефектов в новых кристаллах и обнаружили, что дефекты материнских кристаллов были воспроизведены в дочерних кристаллах, но у дочерних кристаллов также было много дополнительных недостатков. Для того чтобы можно было наблюдать геноподобное поведение, количество наследований этих недостатков должно было превышать количество мутаций в последующих поколениях, но этого не произошло. Таким образом, Кар пришел к выводу, что кристаллы «недостаточно точны, чтобы хранить и передавать информацию от одного поколения к другому».[316]
Железно-серный мир
В 1980-х годах Гюнтер Вехтерсхойзер при поддержке и поддержке Карл Поппер,[317][318][319] постулировал свой железо-серный мир, теорию эволюции предбиотических химических путей как отправную точку в эволюции жизни. Он систематически прослеживает сегодняшнюю биохимию до первичных реакций, которые обеспечивают альтернативные пути к синтезу органических строительных блоков из простых газообразных соединений.
В отличие от классических экспериментов Миллера, которые зависят от внешних источников энергии (имитация молнии, ультрафиолетовое облучение ), «Wächtershäuser systems» имеют встроенный источник энергии: сульфиды железа (железа пирит ) и другие минералы. Энергия, высвобождаемая из редокс реакции этих сульфидов металлов доступны для синтеза органических молекул, и такие системы могли развиться в автокаталитические комплексы, представляющие собой самовоспроизводящиеся, метаболически активные сущности, предшествовавшие известным сегодня формам жизни.[20][312] Эксперименты с такими сульфидами в водной среде при 100 ° C дали относительно небольшой выход дипептиды (От 0,4% до 12,4%) и меньшая доходность трипептиды (0,10%), хотя в тех же условиях дипептиды быстро разрушались.[320]
Некоторые модели отвергают саморепликацию «голого гена», вместо этого постулируя появление примитивного метаболизма, обеспечивающего безопасную среду для более позднего появления репликации РНК. Центральное место в Цикл Кребса (цикл лимонной кислоты) для производства энергии в аэробных организмах, а также для поглощения углекислого газа и ионов водорода при биосинтезе сложных органических химикатов, предполагает, что это была одна из первых частей метаболизма, которая эволюционировала.[290] Соответственно, геохимик Рассел предположил, что «цель жизни — гидрогенизация углекислого газа» (как часть сценария «прежде всего метаболизм», а не «генетика прежде всего»).[321][322] Физик Джереми Инглэнд предположил, что жизнь была неизбежна из общих термодинамических соображений:
… когда группа атомов приводится в движение внешним источником энергии (например, солнцем или химическим топливом) и окружена тепловой ванной (например, океан или атмосфера), она часто постепенно реструктурируется, чтобы рассеивать все больше и больше. энергия. Это может означать, что при определенных условиях материя неумолимо приобретает ключевой физический атрибут, связанный с жизнью.[323][324]
Одно из самых ранних воплощений этой идеи было выдвинуто в 1924 году с представлением Опарина о примитивных самовоспроизводящихся везикулах, которое предшествовало открытию структуры ДНК. Варианты 1980-х и 1990-х годов включают теорию мира железа и серы Вехтерсхойзера и модели, представленные Кристиан де Дюв на основе химии тиоэфиры. Более абстрактные и теоретические аргументы в пользу вероятности возникновения метаболизма без присутствия генов включают математическую модель, введенную Фриман Дайсон в начале 1980-х и Стюарт Кауфман понятие коллективно автокаталитических наборов, обсуждавшееся позже в том же десятилетии.
Оргель резюмировал свой анализ, заявив:
В настоящее время нет причин ожидать, что многоступенчатые циклы, такие как цикл восстановительной лимонной кислоты, будут самоорганизовываться на поверхности FeS / FeS.2 или какой-то другой минерал «.[325]
Возможно, в начале жизни использовался другой тип метаболического пути. Например, вместо цикла восстановительной лимонной кислоты «открытый» ацетил-КоА Путь (еще один из пяти признанных сегодня способов фиксации углекислого газа в природе) был бы совместим с идеей самоорганизации на поверхности сульфида металла. Ключевой фермент этого пути, угарный газ дегидрогеназа /ацетил-КоА-синтаза, содержит смешанные кластеры никель-железо-сера в своих реакционных центрах и катализирует образование ацетил-КоА (аналогично ацетилтиолу) за одну стадию. Однако растет беспокойство по поводу того, что пребиотик тиолированный и тиоэфир соединения термодинамически и кинетически неблагоприятны для накопления в предполагаемых пребиотических условиях (например, в гидротермальных жерлах).[326] Также было предложено, чтобы цистеин и гомоцистеин возможно, отреагировал нитрилы в результате Реакция Стекера, легко образуя каталитические поп-липептиды, достигаемые тиолами.[327]
Гипотеза цинкового мира
Цинковый мир (Zn-мир) теория Мулкиджана[328] является расширением гипотезы пирита Вехтерсхойзера. Вахтерсхойзер основал свою теорию начальных химических процессов, ведущих к информационным молекулам (РНК, пептиды), на регулярной сети электрических зарядов на поверхности пирита, которая, возможно, способствовала первозданному полимеризация притягивая реагенты и располагая их соответствующим образом относительно друг друга.[329] Теория Zn-мира уточняет и дифференцирует дальше.[328][330] Гидротермальные жидкости, богатые H2S, взаимодействуя с холодной первобытной океанской (или «теплым прудом» Дарвина) водой, приводит к осаждению частиц сульфидов металлов. Океанический вентиляционные системы и другие гидротермальные системы имеют зональную структуру, отраженную в древних вулканогенные массивные сульфидные месторождения (VMS) гидротермального происхождения. Они достигают многих километров в диаметре и восходят к Архейский Eon. Наиболее распространены пирит (FeS2), халькопирит (CuFeS2), и сфалерит (ZnS), с добавками галенит (PbS) и алабандит (MnS). ZnS и MnS обладают уникальной способностью накапливать энергию излучения, например от ультрафиолета. Во время соответствующего временного окна происхождения реплицирующихся молекул первичное атмосферное давление было достаточно высоким (> 100 бар, около 100 атмосфер) для осаждения у поверхности Земли, а УФ-излучение было в 10-100 раз более интенсивным, чем сейчас; следовательно, уникальные фотосинтетические свойства, опосредованные ZnS, обеспечили правильные энергетические условия для активизации синтеза информационных и метаболических молекул и выбора фотостабильных азотистых оснований.
Теория Zn-мира была дополнительно дополнена экспериментальными и теоретическими доказательствами ионного строения внутренней части первых прото-клеток до архей, бактерий и протоэукариоты эволюционировал. Арчибальд Макаллум отметили сходство жидкостей организма, таких как кровь и лимфа, с морской водой;[331] однако неорганический состав всех ячеек отличается от состава современной морской воды, что побудило Мулкиджаняна и его коллег реконструировать «инкубаторы» первых ячеек, сочетая геохимический анализ с филогеномный изучение требований к неорганическим ионам универсальных компонентов современных элементов. Авторы приходят к выводу, что повсеместные и, как следует из первичных, белки и функциональные системы демонстрируют сродство и функциональную потребность в K+, Zn2+, Mn2+, и [PO
4]3−
. Геохимическая реконструкция показывает, что ионный состав, способствующий происхождению клеток, не мог существовать в том, что мы сегодня называем морскими условиями, но он совместим с выбросами зон с преобладанием пара в том, что мы сегодня называем внутренними геотермальными системами. При недостатке кислорода CO2— преобладание первичной атмосферы, химический состав водяных конденсатов и испарений вблизи геотермальных полей напоминал бы внутреннюю среду современных ячеек. Следовательно, доклеточные стадии эволюции могли происходить в неглубоких «прудах Дарвина», выстланных пористыми силикатные минералы смешанный с сульфидами металлов и обогащенный калием+, Zn2+, и соединения фосфора.[332][333]
Другие сценарии абиогенеза
Мы определяем сценарий как набор связанных понятий, относящихся к происхождению жизни, которая исследуется или исследовалась. Концепции, относящиеся к миру железо-сера, можно рассматривать как сценарий. Мы рассматриваем некоторые другие сценарии, которые могут частично совпадать со сценариями, описанными выше, или друг с другом.
Химические пути, описанные компьютером
В сентябре 2020 года химики впервые описали возможные химические пути от неживых пребиотических химикатов к сложные биохимические вещества что могло привести к живые организмы, основанный на новой компьютерной программе под названием ALLCHEMY.[334][335]
Гиперцикл
В начале 1970-х Манфред Эйген и Питер Шустер исследовали переходные стадии между молекулярным хаосом и самовоспроизводящимся гиперцикл в супе с пребиотиками.[336] В гиперцикле Информация система хранения (возможно, РНК) производит фермент, который катализирует формирование другой информационной системы, последовательно до тех пор, пока продукт последней не поможет в формировании первой информационной системы. С математической точки зрения гиперциклы могут создавать квазивиды, который посредством естественного отбора вошел в форму дарвиновской эволюции. Толчком к теории гиперциклов стало открытие рибозимы способны катализировать собственные химические реакции. Теория гиперцикла требует существования сложных биохимических веществ, таких как нуклеотиды, которые не образуются в условиях, предложенных экспериментом Миллера-Юри.
Органические пигменты в диссипативных структурах
В своей «Термодинамической теории рассеяния происхождения и эволюции жизни»,[337][55][54] Каро Микаэлиан довел понимание Больцмана и работы Пригожина до их окончательных последствий в отношении происхождения жизни. Эта теория постулирует, что отличительной чертой происхождения и эволюции жизни является микроскопическое диссипативное структурирование органические пигменты и их распространение по всей поверхности Земли.[54] Современная жизнь увеличивает производство энтропии Земли в ее солнечной среде, рассеивая ультрафиолетовый и видимый фотоны в тепло через органические пигменты в воде. Затем это тепло катализирует множество вторичных диссипативных процессов, таких как круговорот воды, океан и ветер токи, ураганы, так далее.[55][56] Михаэлиан утверждает, что если термодинамическая функция жизни сегодня состоит в том, чтобы производить энтропию посредством рассеяния фотонов в органических пигментах, то, вероятно, это была ее функция с самого начала. Оказывается, оба РНК и ДНК когда в водном растворе присутствуют очень сильные поглотители и чрезвычайно быстро рассеивающие ультрафиолетовые лучи в диапазоне длин волн 230–290 нм (УФ-С), который является частью солнечного спектра, который мог проникнуть через пребиотик атмосфера.[338] Фактически, не только РНК и ДНК, но и многие фундаментальные молекулы жизни (общие для всех трех домены жизни) также являются пигментами, поглощающими УФ-С, и многие из них также обладают химическим сродством к РНК и ДНК.[339] Нуклеиновые кислоты Таким образом, могли действовать как молекулы-акцепторы для фотона УФ-С в восторге молекулы донора пигмента антенны, обеспечивая сверхбыстрый канал для рассеивания. Михаэлиан показал, используя формализм нелинейной необратимой термодинамики, что она существовала бы во время Архейский термодинамический императив абиогенного УФ-С фотохимический синтез и распространение этих пигментов по всей поверхности Земли, если они действовали как катализаторы для увеличения рассеивания солнечных фотонов.[340] К концу архея с индуцированным жизнью озон рассеивая УФ-С свет в верхних слоях атмосферы Земли, возникновение совершенно новой жизни, которая не зависела бы от уже существующих сложных метаболических путей, стало бы еще более невероятным, поскольку теперь свободная энергия фотонов, достигающих поверхности Земли, будет иметь было недостаточно для прямой разборки и переделки ковалентные связи. Было высказано предположение, однако, что такие изменения в поверхностном потоке ультрафиолетового излучения из-за геофизических событий, влияющих на атмосферу, могли быть тем, что способствовало развитию сложности в жизни, основанной на существующих метаболических путях, например, во время Кембрийский взрыв[341]
Некоторые из самых сложных проблем, связанных с происхождением жизни, например, отсутствие ферментов. репликация РНК и ДНК,[342] гомохиральность фундаментальных молекул,[343] и происхождение кодирование информации в РНК и ДНК, также найти объяснение в рамках той же диссипативной термодинамики, рассматривая возможное существование связи между первичной репликацией и диссипацией фотонов УФ-С. Михаэлиан предполагает, что ошибочно ожидать описания возникновения, распространения или даже эволюции жизни без чрезмерной ссылки на производство энтропии через диссипацию обобщенного термодинамического потенциала, в частности, преобладающего потока солнечных фотонов.
Белковый амилоид
A new origin-of-life theory based on self-replicating beta-sheet structures has been put forward by Maury in 2009.[344][345] The theory suggest that self-replicating and self-assembling catalytic амилоиды were the first informational polymers in a primitive pre-RNA world. The main arguments for the амилоидная гипотеза is based on the structural stability, autocatalytic and catalytic properties, and evolvability of beta-sheet based informational systems. Such systems are also error correcting[346] и chiroselective.[347]
Колеблющаяся соленость: разбавление и высыхание
Theories of abiogenesis seldom address the caveat raised by Harold Blum:[348] if the key informational elements of life – proto-nucleic acid chains – spontaneously form duplex structures, then there is no way to dissociate them.
Somewhere in this cycle work must be done, which means that free energy must be expended. If the parts assemble themselves on a template spontaneously, work has to be done to take the replica off; or, if the replica comes off the template of its own accord, work must be done to put the parts on in the first place.
The Oparin–Haldane conjecture addresses the formation, but not the dissociation, of nucleic acid polymers and duplexes. However, nucleic acids are unusual because, in the absence of counterions (low salt) to neutralize the high charges on opposing phosphate groups, the nucleic acid duplex dissociates into single chains.[349] Early tides, driven by a close moon, could have generated rapid cycles of dilution (high tide, low salt) and concentration (dry-down at low tide, high salt) that exclusively promoted the replication of nucleic acids[349] through a process dubbed tidal chain reaction (TCR).[350] This theory has been criticized on the grounds that early tides may not have been so rapid,[351] although regression from current values requires an Earth–Moon juxtaposition at around two Ga, for which there is no evidence, and early tides may have been approximately every seven hours.[352] Another critique is that only 2–3% of the Earth’s crust may have been exposed above the sea until late in terrestrial evolution.[353]
The TCR (tidal chain reaction) theory has mechanistic advantages over thermal association/dissociation at deep-sea vents because TCR requires that chain assembly (template-driven polymerization) takes place during the dry-down phase, when precursors are most concentrated, whereas thermal cycling needs polymerization to take place during the cold phase, when the rate of chain assembly is lowest and precursors are likely to be more dilute.
Первый белок, который конденсирует субстрат во время термоциклирования: термосинтез.

Convection cells in fluid placed in a gravity field are selforganizing and enable thermal cycling of the suspended contents in the fluid such as protocells containing protoenzymes that work on thermal cycling.
Emergence of chemiosmotic machinery Today’s bioenergetic process of ферментация is carried out by either the aforementioned citric acid cycle or the Acetyl-CoA pathway, both of which have been connected to the primordial Iron–sulfur world.
In a different approach, the thermosynthesis hypothesis considers the bioenergetic process of хемиосмос, which plays an essential role in клеточное дыхание and photosynthesis, more basal than fermentation: the АТФ-синтаза enzyme, which sustains chemiosmosis, is proposed as the currently extant enzyme most closely related to the first metabolic process.[354][355]
First life needed an energy source to bring about the condensation reaction that yielded the peptide bonds of proteins and the phosphodiester bonds of RNA. In a generalization and thermal variation of the механизм изменения привязки of today’s ATP synthase, the «first protein» would have bound substrates (peptides, phosphate, nucleosides, RNA ‘monomers’) and condensed them to a reaction product that remained bound until it was released after a temperature change by a thermal unfolding. The primordial first protein would therefore have strongly resembled the beta subunits of the ATP synthase alpha/beta subunits of today’s F1 moiety in the FоF1 АТФ-синтаза. Note however that today’s enzymes function during isothermal conditions, whereas the hypothetical first protein worked on and during thermal cycling.
The energy source under the thermosynthesis hypothesis was thermal cycling, the result of suspension of protocells in a конвекция current, as is plausible in a volcanic hot spring; the convection accounts for the self-organization and dissipative structure required in any origin of life model. The still ubiquitous role of thermal cycling in germination and cell division is considered a relic of primordial thermosynthesis.
К фосфорилирование cell membrane lipids, this first protein gave a selective advantage to the lipid protocell that contained the protein. This protein also synthesized a library of many proteins, of which only a minute fraction had thermosynthesis capabilities. As proposed by Dyson,[14] it propagated functionally: it made daughters with similar capabilities, but it did not copy itself. Functioning daughters consisted of different amino acid sequences.
Whereas the iron–sulfur world identifies a circular pathway as the most simple, the thermosynthesis hypothesis does not even invoke a pathway: ATP synthase’s binding change mechanism resembles a physical adsorption process that yields free energy,[356] rather than a regular enzyme’s mechanism, which decreases the free energy.
The described first protein may be simple in the sense that is requires only a short sequence of conserved amino acid residues, a sequent sufficient for the appropriate catalytic cleft. In contrast, it has been claimed that the emergence of cyclic systems of protein catalysts such as required by fermentation is implausible because of the length of many required sequences.[357]
Мир пре-РНК: проблема рибозы и ее обход
It is possible that a different type of nucleic acid, such as peptide nucleic acid, threose nucleic acid или же glycol nucleic acid, was the first to emerge as a self-reproducing molecule, only later replaced by RNA.[358][359] Ларральд и другие., say that
the generally accepted prebiotic synthesis of рибоза, the formose reaction, yields numerous sugars without any selectivity.[360]
and they conclude that their
results suggest that the backbone of the first genetic material could not have contained ribose or other sugars because of their instability.
The ester linkage of ribose and phosphoric acid in RNA is known to be prone to hydrolysis.[361]
Pyrimidine ribonucleosides and their respective nucleotides have been prebiotically synthesized by a sequence of reactions which by-pass the free sugars, and are assembled in a stepwise fashion by using nitrogenous or oxygenous chemistries. Sutherland has demonstrated high yielding routes to cytidine and uridine ribonucleotides built from small 2 and 3 carbon fragments such as гликолевый альдегид, glyceraldehyde или же glyceraldehyde-3-phosphate, цианамид и цианоацетилен. One of the steps in this sequence allows the isolation of энантиочистка ribose aminooxazoline if the enantiomeric excess of glyceraldehyde is 60% or greater.[362] This can be viewed as a prebiotic purification step, where the said compound spontaneously crystallized out from a mixture of the other pentose aminooxazolines. Ribose aminooxazoline can then react with cyanoacetylene in a mild and highly efficient manner to give the alpha cytidine ribonucleotide. Photoanomerization with UV light allows for inversion about the 1′ anomeric centre to give the correct beta стереохимия.[363] In 2009 they showed that the same simple building blocks allow access, via phosphate controlled nucleobase elaboration, to 2′,3′-cyclic pyrimidine nucleotides directly, which are known to be able to полимеризовать into RNA. This paper also highlights the possibility for the photo-sanitization of the pyrimidine-2′,3′-cyclic phosphates.[364]
Структуры РНК
While features of самоорганизация и самовоспроизведение are often considered the hallmark of living systems, there are many instances of abiotic molecules exhibiting such characteristics under proper conditions. Stan Palasek suggested based on a theoretical model that self-assembly of ribonucleic acid (RNA) molecules can occur spontaneously due to physical factors in hydrothermal vents.[365] Вирус self-assembly within host cells has implications for the study of the origin of life,[366] as it lends further credence to the hypothesis that life could have started as self-assembling organic molecules.[367][368]
Вирусное происхождение
Recent evidence for a «virus first» hypothesis, which may support theories of the RNA world, has been suggested.[369][370] One of the difficulties for the study of the origins of viruses is their high rate of mutation; this is particularly the case in RNA retroviruses like HIV.[371] A 2015 study compared protein fold structures across different branches of the tree of life, where researchers can reconstruct the evolutionary histories of the folds and of the organisms whose геномы code for those folds. They argue that protein folds are better markers of ancient events as their three-dimensional structures can be maintained even as the sequences that code for those begin to change.[369] Thus, the viral protein repertoire retain traces of ancient evolutionary history that can be recovered using advanced биоинформатика подходы. Those researchers think that «the prolonged pressure of genome and particle size reduction eventually reduced virocells into modern viruses (identified by the complete loss of cellular makeup), meanwhile other coexisting cellular lineages diversified into modern cells.»[372] The data suggest that viruses originated from ancient cells that co-existed with the ancestors of modern cells. These ancient cells likely contained segmented RNA genomes.[369][373]
A computational model (2015) has shown that virus capsids may have originated in the Мир РНК and that they served as a means of horizontal transfer between replicator communities since these communities could not survive if the number of gene parasites increased, with certain genes being responsible for the formation of these structures and those that favored the survival of self-replicating communities.[374] The displacement of these ancestral genes between cellular organisms could favor the appearance of new viruses during evolution.[375] Viruses retain a replication module inherited from the prebiotic stage since it is absent in cells.[375] So this is evidence that viruses could originate from the RNA world and could also emerge several times in evolution through genetic escape in cells.[375]
Мир РНК

A number of hypotheses of formation of RNA have been put forward. С 1994 г., there were difficulties in the explanation of the abiotic synthesis of the nucleotides cytosine and uracil.[376] Subsequent research has shown possible routes of synthesis; for example, formamide produces all four ribonucleotides and other biological molecules when warmed in the presence of various terrestrial minerals.[226][227] Early cell membranes could have formed spontaneously from proteinoids, which are protein-like molecules produced when amino acid solutions are heated while in the correct concentration of aqueous solution. These are seen to form micro-spheres which are observed to behave similarly to membrane-enclosed compartments. Other possible means of producing more complicated organic molecules include chemical reactions that take place on глина substrates or on the surface of the mineral пирит.
Factors supporting an important role for RNA in early life include its ability to act both to store information and to catalyze chemical reactions (as a ribozyme); its many important roles as an intermediate in the expression of and maintenance of the genetic information (in the form of DNA) in modern organisms; and the ease of chemical synthesis of at least the components of the RNA molecule under the conditions that approximated the early Earth.[377]
Relatively short RNA molecules have been synthesized, capable of replication.[378] Such replicase RNA, which functions as both code and catalyst provides its own template upon which copying can occur. Szostak has shown that certain catalytic RNAs can join smaller RNA sequences together, creating the potential for self-replication. If these conditions were present, Darwinian natural selection would favour the proliferation of such autocatalytic sets, to which further functionalities could be added.[379] Such autocatalytic systems of RNA capable of self-sustained replication have been identified.[380] The RNA replication systems, which include two ribozymes that catalyze each other’s synthesis, showed a doubling time of the product of about one hour, and were subject to natural selection under the conditions that existed in the experiment.[381] In evolutionary competition experiments, this led to the emergence of new systems which replicated more efficiently.[18] This was the first demonstration of evolutionary adaptation occurring in a molecular genetic system.[381]
Depending on the definition, life started when RNA chains began to self-replicate, initiating the three mechanisms of Darwinian selection: наследственность, variation of type, and differential reproductive output. The fitness of an RNA replicator (its per capita rate of increase) would likely be a function of its intrinsic adaptive capacities, determined by its nucleotide sequence, and the availability of resources.[382][383] The three primary adaptive capacities may have been: (1) replication with moderate fidelity, giving rise to both heritability while allowing variation of type, (2) resistance to decay, and (3) acquisition of process resources.[382][383] These capacities would have functioned by means of the folded configurations of the RNA replicators resulting from their nucleotide sequences.
Эксперименты по происхождению жизни

Дж. Крейг Вентер
Both Eigen and Соль Шпигельман demonstrated that evolution, including replication, variation, and естественный отбор, can occur in populations of molecules as well as in organisms.[49] Following on from chemical evolution came the initiation of биологическая эволюция, which led to the first cells.[49] No one has yet synthesized a «протоклетка » using simple components with the necessary properties of life (the so-called «bottom-up-approach «). Without such a proof-of-principle, explanations have tended to focus on хемосинтез.[384] However, some researchers work in this field, notably Steen Rasmussen and Szostak.
Others have argued that a «нисходящий подход » is more feasible, starting with simple forms of current life. Spiegelman took advantage of natural selection to synthesize the Spiegelman Monster, which had a genome with just 218 нуклеотид bases, having deconstructively evolved from a 4500-base bacterial RNA. Eigen built on Spiegelman’s work and produced a similar system further degraded to just 48 or 54 nucleotides—the minimum required for the binding of the replication enzyme.[385] Крейг Вентер и другие на Институт Дж. Крейга Вентера engineered existing prokaryotic cells with progressively fewer genes, attempting to discern at which point the most minimal requirements for life are reached.[386][387][388]
In October 2018, researchers at Университет Макмастера announced the development of a new technology, called a Симулятор планеты, to help study the происхождение жизни на планете земной шар и дальше.[389][390][391][392] It consists of a sophisticated climate chamber to study how the building blocks of life were assembled and how these prebiotic molecules transitioned into self-replicating RNA molecules.[389]
Смотрите также
- Антропный принцип – Philosophical premise that all scientific observations presuppose a universe compatible with the emergence of sentient organisms that make those observations
- Искусственная ячейка
- Искусственная жизнь — Область исследования, в которой исследователи изучают системы, связанные с естественной жизнью, ее процессами и ее эволюцией, с помощью моделирования.
- Bathybius haeckelii
- Энтропия и жизнь
- Formamide-based prebiotic chemistry
- GADV-protein world hypothesis
- Гемолитин – Protein claimed to be of extraterrestrial origin
- Гипотетические типы биохимии – Possible alternative biochemicals used by life forms
- Mediocrity principle
- Связь для науки об экзопланетах – Dedicated to the search for life on exoplanets
- Ноогенез – Emergence and evolution of intelligence
- Планетарная обитаемость — Степень, в которой планета пригодна для жизни, какой мы ее знаем.
- Protocell – Lipid globule proposed as a precursor of living cells
- Гипотеза редкой земли – Hypothesis that complex extraterrestrial life is improbable and extremely rare
- Теневая биосфера – A hypothetical microbial biosphere of Earth that would use radically different biochemical and molecular processes from that of currently known life
- Толин – Class of molecules formed by ultraviolet irradiation of organic compounds
Рекомендации
Сноски
- ^ Also occasionally called biopoiesis (Bernal, 1960, p. 30)
- ^ The reactions are:
- FeS + H2S → FeS2 + 2H+ + 2e−
- FeS + H2S + CO2 → FeS2 + HCOOH
- ^ The reactions are:
Reaction 1: Fayalite + water → magnetite + aqueous silica + hydrogen- 3Fe2SiO4 + 2H2O → 2Fe3О4 + 3SiO2 + 2H2
Reaction 2: Forsterite + aqueous silica → serpentine
- 3Mg2SiO4 + SiO2 + 4H2O → 2Mg3Si2О5(ОЙ)4
Reaction 3: Forsterite + water → serpentine + brucite
- 2Mg2SiO4 + 3H2O → Mg3Si2О5(ОЙ)4 + Mg (OH)2
Reaction 3 describes the hydration of olivine with water only to yield змеевик and Mg(OH)2 (брусит ). Serpentine is stable at high pH in the presence of brucite like calcium silicate hydrate, (C-S-H ) phases formed along with портландит (Са (ОН)2) in hardened портландцемент paste after the hydration of belite (Ca2SiO4), the artificial calcium equivalent of forsterite.Analogy of reaction 3 with belite hydration in ordinary Portland cement: Belite + water → C-S-H phase + portlandite
- 2 Ca2SiO4 + 4 часа2O → 3 CaO · 2 SiO2 · 3 H2O + Ca(OH)2
Цитаты
- ^ а б c d е Додд, Мэтью С .; Папино, Доминик; Гренн, Тор; Slack, Джон Ф .; Риттнер, Мартин; Пирайно, Франко; О’Нил, Джонатан; Little, Crispin T.S. (1 марта 2017 г.). «Доказательства ранней жизни в осадках старейших гидротермальных источников Земли». Природа. 543 (7643): 60–64. Bibcode:2017Натура.543 … 60Д. Дои:10.1038 / природа21377. PMID 28252057. В архиве из оригинала 8 сентября 2017 г.. Получено 2 марта 2017.
- ^ а б c Циммер, Карл (1 марта 2017 г.). «Ученые говорят, что окаменелости канадских бактерий могут быть самыми древними на Земле». Нью-Йорк Таймс. В архиве из оригинала 2 марта 2017 г.. Получено 2 марта 2017.
- ^ Oparin, Aleksandr Ivanovich (1938). Происхождение жизни. Phoenix Edition Series. Translated by Morgulis, Sergius (2 ed.). Mineola, New York: Courier Corporation (published 2003). ISBN 978-0486495224. Получено 16 июн 2018.
- ^ а б Peretó, Juli (2005). «Controversies on the origin of life» (PDF). Международная микробиология. 8 (1): 23–31. PMID 15906258. Архивировано из оригинал (PDF) 24 августа 2015 г.. Получено 1 июня 2015.
Ever since the historical contributions by Aleksandr I. Oparin, in the 1920s, the intellectual challenge of the origin of life enigma has unfolded based on the assumption that life originated on Earth through physicochemical processes that can be supposed, comprehended, and simulated; that is, there were neither miracles nor spontaneous generations.
- ^ Сравнивать: Scharf, Caleb; и другие. (18 December 2015). «A Strategy for Origins of Life Research». Астробиология. 15 (12): 1031–1042. Bibcode:2015AsBio..15.1031S. Дои:10.1089/ast.2015.1113. ЧВК 4683543. PMID 26684503.
What do we mean by the origins of life (OoL)? […] Since the early 20th century the phrase OoL has been used to refer to the events that occurred during the transition from non-living to living systems on Earth, i.e., the origin of terrestrial biology (Oparin, 1924; Haldane, 1929). The term has largely replaced earlier concepts such as abiogenesis (Kamminga, 1980; Fry, 2000).
- ^ Oparin 1953, п. vi
- ^ Warmflash, David; Warmflash, Benjamin (November 2005). «Did Life Come from Another World?». Scientific American. 293 (5): 64–71. Bibcode:2005SciAm.293e..64W. Дои:10.1038/scientificamerican1105-64. PMID 16318028.
According to the conventional hypothesis, the earliest living cells emerged as a result of chemical evolution on our planet billions of years ago in a process called abiogenesis.
- ^ Yarus 2010, п. 47
- ^ Witzany, Guenther (2016). «Crucial steps to life: From chemical reactions to code using agents» (PDF). BioSystems. 140: 49–57. Дои:10.1016/j.biosystems.2015.12.007. PMID 26723230.
- ^ Хауэлл, Элизабет (8 декабря 2014 г.). «Как жизнь стала сложной и могло ли это произойти за пределами Земли?». Журнал Astrobiology. Получено 14 февраля 2018.
- ^ Tirard, Stephane (20 April 2015). Abiogenesis – Definition. Энциклопедия астробиологии. п. 1. Дои:10.1007/978-3-642-27833-4_2-4. ISBN 978-3-642-27833-4.
Thomas Huxley (1825–1895) used the term abiogenesis in an important text published in 1870. He strictly made the difference between spontaneous generation, which he did not accept, and the possibility of the evolution of matter from inert to living, without any influence of life. […] Since the end of the nineteenth century, evolutive abiogenesis means increasing complexity and evolution of matter from inert to living state in the abiotic context of evolution of primitive Earth.
- ^ Levinson, Gene (2020). Rethinking evolution: the revolution that’s hiding in plain sight. World Scientific. ISBN 978-1786347268.
- ^ Voet & Voet 2004, п. 29
- ^ а б Dyson 1999
- ^ Davies, Paul (1998). The Fifth Miracle, Search for the origin and meaning of life. Пингвин.[страница нужна ]
- ^ Уорд, Питер; Kirschvink, Joe (2015). A New History of Life: the radical discoveries about the origins and evolution of life on earth. Bloomsbury Press. С. 39–40. ISBN 978-1608199105.
- ^ а б *Copley, Shelley D.; Смит, Эрик; Morowitz, Harold J. (Декабрь 2007 г.). «The origin of the RNA world: Co-evolution of genes and metabolism» (PDF). Bioorganic Chemistry. 35 (6): 430–443. Дои:10.1016/j.bioorg.2007.08.001. PMID 17897696. В архиве (PDF) из оригинала 5 сентября 2013 г.. Получено 8 июн 2015.
The proposal that life on Earth arose from an RNA world is widely accepted.
- Orgel, Leslie E. (Апрель 2003 г.). «Some consequences of the RNA world hypothesis». Истоки жизни и эволюция биосферы. 33 (2): 211–218. Bibcode:2003OLEB…33..211O. Дои:10.1023/A:1024616317965. PMID 12967268. S2CID 32779859.
It now seems very likely that our familiar DNA/RNA/protein world was preceded by an RNA world…
- Robertson & Joyce 2012: «There is now strong evidence indicating that an RNA World did indeed exist before DNA- and protein-based life.»
- Neveu, Kim & Benner 2013: «[The RNA world’s existence] has broad support within the community today.»
- Orgel, Leslie E. (Апрель 2003 г.). «Some consequences of the RNA world hypothesis». Истоки жизни и эволюция биосферы. 33 (2): 211–218. Bibcode:2003OLEB…33..211O. Дои:10.1023/A:1024616317965. PMID 12967268. S2CID 32779859.
- ^ а б c d Robertson, Michael P.; Joyce, Gerald F. (Май 2012 г.). «The origins of the RNA world». Перспективы Колд-Спринг-Харбор в биологии. 4 (5): a003608. Дои:10.1101/cshperspect.a003608. ЧВК 3331698. PMID 20739415.
- ^ а б c d Cech, Thomas R. (Июль 2012 г.). «The RNA Worlds in Context». Перспективы Колд-Спринг-Харбор в биологии. 4 (7): a006742. Дои:10.1101/cshperspect.a006742. ЧВК 3385955. PMID 21441585.
- ^ а б c Keller, Markus A.; Turchyn, Alexandra V.; Ralser, Markus (25 March 2014). «Non‐enzymatic glycolysis and pentose phosphate pathway‐like reactions in a plausible Archean ocean». Молекулярная системная биология. 10 (725): 725. Дои:10.1002/msb.20145228. ЧВК 4023395. PMID 24771084.
- ^ Rampelotto, Pabulo Henrique (26 April 2010). Panspermia: A Promising Field of Research (PDF). Astrobiology Science Conference 2010. Хьюстон, Техас: Лунно-планетарный институт. п. 5224. Bibcode:2010LPICo1538.5224R. В архиве (PDF) из оригинала 27 марта 2016 г.. Получено 3 декабря 2014. Conference held at League City, TX
- ^ Berera, Arjun (6 November 2017). «Space dust collisions as a planetary escape mechanism». Астробиология. 17 (12): 1274–1282. arXiv:1711.01895. Bibcode:2017AsBio..17.1274B. Дои:10.1089/ast.2017.1662. PMID 29148823. S2CID 126012488.
- ^ Chan, Queenie H.S. (10 января 2018 г.). «Органическое вещество в кристаллах внеземных водоносных солей». Достижения науки. 4 (1, eaao3521): eaao3521. Bibcode:2018SciA …. 4O3521C. Дои:10.1126 / sciadv.aao3521. ЧВК 5770164. PMID 29349297.
- ^ а б c Эренфройнд, Паскаль; Cami, Jan (December 2010). «Cosmic carbon chemistry: from the interstellar medium to the early Earth». Перспективы Колд-Спринг-Харбор в биологии. 2 (12): a002097. Дои:10.1101/cshperspect.a002097. ЧВК 2982172. PMID 20554702.
- ^ Perkins, Sid (8 April 2015). «Organic molecules found circling nearby star». Наука (News). Вашингтон, округ Колумбия: Американская ассоциация развития науки. Получено 2 июн 2015.
- ^ King, Anthony (14 April 2015). «Chemicals formed on meteorites may have started life on Earth». Chemistry World (News). Лондон: Королевское химическое общество. В архиве из оригинала 17 апреля 2015 г.. Получено 17 апреля 2015.
- ^ Saladino, Raffaele; Carota, Eleonora; Botta, Giorgia; и другие. (13 апреля 2015 г.). «Meteorite-catalyzed syntheses of nucleosides and of other prebiotic compounds from formamide under proton irradiation». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 112 (21): E2746–E2755. Bibcode:2015PNAS..112E2746S. Дои:10.1073/pnas.1422225112. ЧВК 4450408. PMID 25870268.
- ^ Graham, Robert W. (February 1990). «Внеземная жизнь во Вселенной» (PDF) (NASA Technical Memorandum 102363). Исследовательский центр Льюиса, Cleveland, Ohio: НАСА. В архиве (PDF) из оригинала от 3 сентября 2014 г.. Получено 2 июн 2015.
- ^ Altermann 2009, п. xvii
- ^ «Возраст Земли». Геологическая служба США. 9 июля 2007 г. В архиве с оригинала 23 декабря 2005 г.. Получено 10 января 2006.
- ^ Dalrymple 2001, pp. 205–221
- ^ Манхеса, Жерар; Allègre, Claude J.; Дюпреа, Бернар; Hamelin, Bruno (May 1980). «Свинцовые изотопные исследования базовых-ультраосновных слоистых комплексов: предположения о возрасте Земли и характеристиках примитивной мантии». Письма по науке о Земле и планетах. 47 (3): 370–382. Bibcode:1980E и PSL..47..370M. Дои:10.1016 / 0012-821X (80) 90024-2.
- ^ а б Schopf, J. William; Кудрявцев Анатолий Б .; Czaja, Andrew D .; Трипати, Абхишек Б. (5 октября 2007 г.). «Evidence of Archean life: Stromatolites and microfossils». Докембрийские исследования. 158 (3–4): 141–155. Bibcode:2007Пред..158..141С. Дои:10.1016 / j.precamres.2007.04.009.
- ^ а б Шопф, Дж. Уильям (29 июня 2006 г.). «Fossil evidence of Archaean life». Философские труды Королевского общества B. 361 (1470): 869–885. Дои:10.1098/rstb.2006.1834. ЧВК 1578735. PMID 16754604.
- ^ а б Raven & Johnson 2002, п. 68
- ^ Staff (9 May 2017). «Oldest evidence of life on land found in 3.48-billion-year-old Australian rocks». Phys.org. В архиве с оригинала 10 мая 2017 г.. Получено 13 мая 2017.
- ^ Джокич, Тара; Ван Кранендонк, Мартин Дж .; Кэмпбелл, Кэтлин А .; Уолтер, Малкольм Р .; Уорд, Колин Р. (9 мая 2017 г.). «Earliest signs of life on land preserved in ca. 3.5 Gao hot spring deposits». Nature Communications. 8: 15263. Bibcode:2017NatCo…815263D. Дои:10.1038 / ncomms15263. ЧВК 5436104. PMID 28486437.
- ^ Шопф, Дж. Уильям; Kitajima, Kouki; Spicuzza, Michael J.; Kudryavtsev, Anatolly B.; Valley, John W. (2017). «SIMS analyses of the oldest known assemblage of microfossils document their taxon-correlated carbon isotope compositions». PNAS. 115 (1): 53–58. Bibcode:2018PNAS..115…53S. Дои:10.1073/pnas.1718063115. ЧВК 5776830. PMID 29255053.
- ^ Tyrell, Kelly April (18 December 2017). «Oldest fossils ever found show life on Earth began before 3.5 billion years ago». Университет Висконсин-Мэдисон. Получено 18 декабря 2017.
- ^ Гош, Паллаб (1 марта 2017 г.). «Earliest evidence of life on Earth found». Новости BBC. Новости BBC. В архиве из оригинала 2 марта 2017 г.. Получено 2 марта 2017.
- ^ а б Данэм, Уилл (1 марта 2017 г.). «Канадские окаменелости, похожие на бактерии, называют древнейшими свидетельствами жизни». Рейтер. В архиве из оригинала 2 марта 2017 г.. Получено 1 марта 2017.
- ^ «Researchers uncover ‘direct evidence’ of life on Earth 4 billion years ago». Deutsche Welle. Получено 5 марта 2017.
- ^ а б c d «NASA Astrobiology Strategy» (PDF). НАСА. 2015. Архивировано с оригинал (PDF) 22 декабря 2016 г.. Получено 24 сентября 2017.
- ^ Guthrie, W. K. C. (1957). In the beginning: Some Greek views on the origins of life and the early state of man. Метуэн, Лондон.
- ^ Simon, Michael A. (1971). The Matter of Life (1-е изд.). Нью-Хейвен и Лондон: Издательство Йельского университета.
- ^ Ladyman, J.; Lambert, J.; Weisner, K.B. What is a Complex System? Евро. J. Philos. Sci. 2013, 3,33–67.
- ^ Esposito, M., Lindenberg, K., & Van den Broeck, C. (2010). Entropy production as correlation between system and reservoir. New Journal of Physics, 12(1), 013013.
- ^ Bernal 1967, п. 143
- ^ а б c d е ж грамм час я j Follmann, Hartmut; Brownson, Carol (November 2009). «Darwin’s warm little pond revisited: from molecules to the origin of life». Naturwissenschaften. 96 (11): 1265–1292. Bibcode:2009NW…..96.1265F. Дои:10.1007/s00114-009-0602-1. PMID 19760276. S2CID 23259886.
- ^ Gora, Evan M.; Burchfield, Jeffrey C.; Muller‐Landau, Helene C.; Bitzer, Phillip M.; Yanoviak, Stephen P. (2020). «Pantropical geography of lightning-caused disturbance and its implications for tropical forests». Биология глобальных изменений. н / д (n/a): 5017–5026. Bibcode:2020GCBio..26.5017G. Дои:10.1111/gcb.15227. ISSN 1365-2486. PMID 32564481.
- ^ Kalson, Natan-Haim; Furman, David; Zeiri, Yehuda (2017). «Cavitation-Induced Synthesis of Biogenic Molecules on Primordial Earth». ACS Central Science. 3 (9): 1041–1049. Дои:10.1021/acscentsci.7b00325. ЧВК 5620973. PMID 28979946.
- ^ Haken, Hermann (1978). Синергетика. Введение. Берлин: Springer.
- ^ Haken, Hermann (1978). Синергетика. Введение. Springer.
- ^ а б c Michaelian, Karo (2017). «Microscopic dissipative structuring and proliferation at the origin of life». Гелион. 3 (10): e00424. Дои:10.1016/j.heliyon.2017.e00424. ЧВК 5647473. PMID 29062973.
- ^ а б c Michaelian, K (2011). «Thermodynamic dissipation theory for the origin of life». Earth System Dynamics. 2 (1): 37–51. arXiv:0907.0042. Bibcode:2011ESD…..2…37M. Дои:10.5194/esd-2-37-2011. S2CID 14574109.
- ^ а б Michaelian, K (2012). «HESS Opinions ‘Biological catalysis of the hydrological cycle: Life’s thermodynamic function‘«. Гидрология и науки о Земле. 16 (8): 2629–2645. arXiv:0907.0040. Bibcode:2012HESS…16.2629M. Дои:10.5194/hess-16-2629-2012.
- ^ Boltzmann, L. (1886) The Second Law of Thermodynamics, in: Ludwig Boltzmann: Theoretical physics and Selected writings, edited by: McGinness, B., D. Reidel, Dordrecht, The Netherlands, 1974.
- ^ Schrödinger, Erwin (1944) What is Life? The Physical Aspect of the Living Cell. Издательство Кембриджского университета
- ^ Onsager, L. (1931) Reciprocal Relations in Irreversible Processes I and II, Phys. Ред. 37, 405; 38, 2265 (1931)
- ^ Prigogine, I. (1967) An Introduction to the Thermodynamics of Irreversible Processes, Wiley, New York
- ^ Dewar, R; Juretić, D.; Županović, P. (2006). «The functional design of the rotary enzyme ATP synthase is consistent with maximum entropy production». Chem. Phys. Латыш. 430 (1): 177–182. Bibcode:2006CPL…430..177D. Дои:10.1016/j.cplett.2006.08.095.
- ^ Unrean, P., Srienc, F. (2011) Metabolic networks evolve towards states of maximum entropy production, Metabolic Engineering 13, 666–673.
- ^ Zotin, A.I. (1984) «Bioenergetic trends of evolutionary progress of organisms», in: Thermodynamics and regulation of biological processes Lamprecht, I. and Zotin, A.I. (eds.), De Gruyter, Berlin, pp. 451–458.
- ^ Schneider, E.D.; Kay, J.J. (1994). «Life as a Manifestation of the Second Law of Thermodynamics». Математическое и компьютерное моделирование. 19 (6–8): 25–48. CiteSeerX 10.1.1.36.8381. Дои:10.1016/0895-7177(94)90188-0.
- ^ Michaelian, K. (2005). «Thermodynamic stability of ecosystems». Журнал теоретической биологии. 237 (3): 323–335. Bibcode:2004APS..MAR.P9015M. Дои:10.1016/j.jtbi.2005.04.019. PMID 15978624.
- ^ Trifonov, Edward N. (17 March 2011). «Vocabulary of Definitions of Life Suggests a Definition». Journal of Biomolecular Structure and Dynamics. 29 (2): 259–266. Дои:10.1080/073911011010524992. Получено 15 декабря 2020.
- ^ Gould, James L.; Keeton, William T. (1996). Биологические науки (6 изд.). Нью-Йорк: W.W. Нортон.
- ^ Campbell, Neil A.; Reece, Jane B. (2005). Биология (7-е изд.). Sn Feancisco: Benjamin.
- ^ Casti, John L. (1989). Paradigms lost. Images of man in the mirror of science. Нью-Йорк: Морроу. Bibcode:1989plim.book…..C.
- ^ Шульце-Макух, Дирк; Irwin, Louis N. (2018). Life in the Universe. Expectations and Constraints (3-е изд.). Нью-Йорк: Спрингер.
- ^ Lehninger, Albert L. (1970). Биохимия. The Molecular Basis of Cell Structure and Function. Нью-Йорк: стоит. п. 313.
- ^ Anthonie W.J. Muller (1995). «Were the first organisms heat engines? A new model for biogenesis and the early evolution of biological energy conversion». Прогресс в биофизике и молекулярной биологии. 63 (2): 193–231. Дои:10.1016/0079-6107(95)00004-7. PMID 7542789.
- ^ Anthonie W.J. Muller and Dirk Schulze-Makuch (2006). «Thermal energy and the origin of life». Истоки жизни и эволюция биосфер. 36 (2): 77–189. Bibcode:2006OLEB…36..177M. Дои:10.1007/s11084-005-9003-4. PMID 16642267. S2CID 22179552.
- ^ Wimberly, Brian T.; Brodersen, Ditlev E.; Clemons, William M. Jr.; и другие. (21 сентября 2000 г.). «Structure of the 30S ribosomal subunit». Природа. 407 (6802): 327–339. Bibcode:2000Natur.407..327W. Дои:10.1038/35030006. PMID 11014182. S2CID 4419944.
- ^ Zimmer, Carl (25 September 2014). «A Tiny Emissary From the Ancient Past». Нью-Йорк Таймс. Нью-Йорк. В архиве из оригинала 27 сентября 2014 г.. Получено 26 сентября 2014.
- ^ Уэйд, Николас (4 May 2015). «Making Sense of the Chemistry That Led to Life on Earth». Нью-Йорк Таймс. Нью-Йорк. В архиве из оригинала 9 июля 2017 г.. Получено 10 мая 2015.
- ^ Benner, S.A.; Bell, E.A.; Biondi, E.; Brasser, R.; Carell, T.; Kim, H.-J.; Mojzsis, S.J.; Omran, A.; Pasek, M.A.; Trail, D. (2020). «When Did Life Likely Emerge on Earth in an RNA‐First Process?». ChemSystemsChem. 2 (2). Дои:10.1002/syst.201900035.
- ^ Yarus, Michael (April 2011). «Getting Past the RNA World: The Initial Darwinian Ancestor». Перспективы Колд-Спринг-Харбор в биологии. 3 (4): a003590. Дои:10.1101/cshperspect.a003590. ЧВК 3062219. PMID 20719875.
- ^ Fox, George.E. (9 июня 2010 г.). «Origin and evolution of the ribosome». Перспективы Колд-Спринг-Харбор в биологии. 2 (9(a003483)): a003483. Дои:10.1101/cshperspect.a003483. ЧВК 2926754. PMID 20534711.
- ^ Neveu, Marc; Kim, Hyo-Joong; Benner, Steven A. (22 April 2013). «The ‘Strong’ RNA World Hypothesis: Fifty Years Old». Астробиология. 13 (4): 391–403. Bibcode:2013AsBio..13..391N. Дои:10.1089/ast.2012.0868. PMID 23551238.
- ^ Гилберт, Уолтер (20 February 1986). «Origin of life: The RNA world». Природа. 319 (6055): 618. Bibcode:1986Natur.319..618G. Дои:10.1038/319618a0. S2CID 8026658.
- ^ Gough, Evan (10 March 2020). «Life Could be Common Across the Universe, Just Not in Our Region». Вселенная сегодня. Получено 15 марта 2020.
- ^ Totani, Tomonori (3 February 2020). «Emergence of life in an inflationary universe». Научные отчеты. 10 (1671): 1671. arXiv:1911.08092. Bibcode:2020NatSR..10.1671T. Дои:10.1038/s41598-020-58060-0. ЧВК 6997386. PMID 32015390.
- ^ Бун, Дэвид Р .; Castenholz, Richard W.; Гаррити, Джордж М., ред. (2001). В Археи and the Deeply Branching and Phototrophic Бактерии. Руководство Берджи по систематической бактериологии. Springer. ISBN 978-0-387-21609-6. В архиве from the original on 25 December 2014.[страница нужна ]
- ^ Woese CR, Fox GE (1977). «Phylogenetic structure of the prokaryotic domain: the primary kingdoms». Proc Natl Acad Sci U S A. 74 (11): 5088–5090. Bibcode:1977PNAS … 74.5088W. Дои:10.1073 / пнас.74.11.5088. ЧВК 432104. PMID 270744.
- ^ Valas RE, Bourne PE (2011). «The origin of a derived superkingdom: how a gram-positive bacterium crossed the desert to become an archaeon». Биология Директ. 6: 16. Дои:10.1186/1745-6150-6-16. ЧВК 3056875. PMID 21356104.
- ^ Cavalier-Smith T (2006). «Rooting the tree of life by transition analyses». Биология Директ. 1: 19. Дои:10.1186/1745-6150-1-19. ЧВК 1586193. PMID 16834776.
- ^ Ward, Peter Douglas (2005). Life as We Do Not Know it: The NASA Search for (and Synthesis Of) Alien Life. Viking Books. ISBN 978-0670034581.
- ^ Уэйд, Николас (25 July 2016). «Meet Luca, the Ancestor of All Living Things». Нью-Йорк Таймс. В архиве из оригинала от 28 июля 2016 г.
- ^ Weiss, M.C.; Sousa, F.L.; Mrnjavac, N.; Neukirchen, S.; Roettger, M.; Nelson-Sathi, S.; Martin, W.F. (2016). «Физиология и среда обитания последнего универсального общего предка». Природная микробиология. 1 (9): 16116. Дои:10.1038/NMICROBIOL.2016.116. PMID 27562259. S2CID 2997255.
- ^ Nature Vol 535, 28 July 2016,»Early Life Liked it Hot», p.468
- ^ Noller, Harry F. (Апрель 2012 г.). «Evolution of protein synthesis from an RNA world». Перспективы Колд-Спринг-Харбор в биологии. 4 (4): a003681. Дои:10.1101/cshperspect.a003681. ЧВК 3312679. PMID 20610545.
- ^ Koonin, Eugene V. (31 May 2007). «The cosmological model of eternal inflation and the transition from chance to biological evolution in the history of life». Биология Директ. 2: 15. Дои:10.1186/1745-6150-2-15. ЧВК 1892545. PMID 17540027.
- ^ Hoffmann, Geoffrey W. (25 июня 1974 г.). «On the origin of the genetic code and the stability of the translation apparatus». Журнал молекулярной биологии. 86 (2): 349–362. Дои:10.1016/0022-2836(74)90024-2. PMID 4414916.
- ^ Orgel, Leslie E. (April 1963). «The Maintenance of the Accuracy of Protein Synthesis and its Relevance to Ageing». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 49 (4): 517–521. Bibcode:1963PNAS…49..517O. Дои:10.1073/pnas.49.4.517. ЧВК 299893. PMID 13940312.
- ^ Hoffmann, Geoffrey W. (October 1975). «The Stochastic Theory of the Origin of the Genetic Code». Ежегодный обзор физической химии. 26: 123–144. Bibcode:1975ARPC…26..123H. Дои:10.1146/annurev.pc.26.100175.001011.
- ^ Chaichian, Rojas & Tureanu 2014, pp. 353–364
- ^ Plasson, Raphaël; Kondepudi, Dilip K.; Bersini, Hugues; и другие. (Август 2007 г.). «Emergence of homochirality in far-from-equilibrium systems: Mechanisms and role in prebiotic chemistry». Хиральность. 19 (8): 589–600. Дои:10.1002/chir.20440. PMID 17559107. «Специальный выпуск: материалы восемнадцатого Международного симпозиума по хиральности (ISCD-18), Пусан, Корея, 2006 г.»
- ^ Джафарпур, Фаршид; Бьянкалани, Томмазо; Голденфельд, Найджел (2017). «Вызванное шумом нарушение симметрии вдали от равновесия и появление биологической гомохиральности» (PDF). Физический обзор E. 95 (3): 032407. Bibcode:2017PhRvE..95c2407J. Дои:10.1103 / PhysRevE.95.032407. PMID 28415353.
- ^ Джафарпур, Фаршид; Бьянкалани, Томмазо; Голденфельд, Найджел (2015). «Шум-индуцированный механизм биологической гомохиральности саморепликаторов ранней жизни». Письма с физическими проверками. 115 (15): 158101. arXiv:1507.00044. Bibcode:2015PhRvL.115o8101J. Дои:10.1103 / PhysRevLett.115.158101. PMID 26550754. S2CID 9775791.
- ^ Франк, F.C. (1953). «О спонтанном асимметричном синтезе». Biochimica et Biophysica Acta. 11 (4): 459–463. Дои:10.1016/0006-3002(53)90082-1. PMID 13105666.
- ^ Кларк, Стюарт (Июль – август 1999 г.). «Поляризованный звездный свет и рука жизни». Американский ученый. 87 (4): 336. Bibcode:1999AmSci..87..336C. Дои:10.1511/1999.4.336.
- ^ Шибата, Таканори; Мориока, Хироши; Хаясе, Тадакацу; и другие. (17 января 1996 г.). «Высокоэнантиоселективное каталитическое асимметричное автоматическое размножение хирального пиримидилового спирта». Журнал Американского химического общества. 118 (2): 471–472. Дои:10.1021 / ja953066g.
- ^ Соай, Кенсо; Сато, Итару; Сибата, Таканори (2001). «Асимметричный автокатализ и происхождение хиральной гомогенности в органических соединениях». Химический рекорд. 1 (4): 321–332. Дои:10.1002 / tcr.1017. PMID 11893072.
- ^ Hazen 2005, п. 184
- ^ Мейерхенрих, Уве (2008). Аминокислоты и асимметрия жизни, пойманная в процессе формирования. Берлин: Springer. С. 76–79. ISBN 978-3540768869.
- ^ Маллен, Лесли (5 сентября 2005 г.). «Строим жизнь из звездных вещей». Журнал Astrobiology. В архиве из оригинала 14 июля 2015 г.. Получено 15 июн 2015.
- ^ Раби, Пассан (6 июля 2020 г.). «Астрономы нашли источник жизни во Вселенной». Обратный. Получено 7 июля 2020.
- ^ Мариго, Паола; и другие. (6 июля 2020 г.). «Формирование углеродных звезд через немонотонное соотношение начальной и конечной масс». Природа Астрономия. 152 (11): 1102–1110. arXiv:2007.04163. Bibcode:2020НатАс … 4,1 102 млн. Дои:10.1038 / с41550-020-1132-1. S2CID 220403402. Получено 7 июля 2020.
- ^ «WMAP — Жизнь во Вселенной».
- ^ Формирование солнечных систем: теория солнечных туманностей. Массачусетский университет в Амхерсте, факультет астрономии. Доступ 27 сентября 2019 г.
- ^ Кастинг, Джеймс Ф. (12 февраля 1993 г.). «Ранняя атмосфера Земли» (PDF). Наука. 259 (5097): 920–926. Bibcode:1993Наука … 259..920K. Дои:10.1126 / science.11536547. PMID 11536547. S2CID 21134564. Архивировано из оригинал (PDF) 10 октября 2015 г.. Получено 28 июля 2015.
- ^ Фесенков 1959, п. 9
- ^ Морс, Джон (сентябрь 1998 г.). «Геохимия карбонатов Гадийского океана». Водная геохимия. 4 (3/4): 301–319. Bibcode:1998MinM … 62.1027M. Дои:10.1023 / А: 1009632230875. S2CID 129616933.
- ^ Морс, Джон В .; Маккензи, Фред Т. (1998). «Геохимия карбонатов Гадийского океана». Водная геохимия. 4 (3–4): 301–319. Bibcode:1998MinM … 62.1027M. Дои:10.1023 / А: 1009632230875. S2CID 129616933.
- ^ Уайльд, Саймон А .; Вэлли, Джон В .; Пек, Уильям Х .; Грэм, Колин М. (11 января 2001 г.). «Доказательства существования континентальной коры и океанов на Земле 4,4 млрд лет назад по детритовым цирконам» (PDF). Природа. 409 (6817): 175–178. Bibcode:2001 Натур 409..175 Вт. Дои:10.1038/35051550. PMID 11196637. S2CID 4319774. В архиве (PDF) из оригинала 5 июня 2015 г.. Получено 3 июн 2015.
- ^ Розинг, Миник Т .; Берд, Деннис К .; Сон, Норман Х .; и другие. (22 марта 2006 г.). «Возникновение континентов — очерк геологических последствий фотосинтеза». Палеогеография, палеоклиматология, палеоэкология. 232 (2–4): 99–113. Bibcode:2006ППП … 232 … 99р. Дои:10.1016 / j.palaeo.2006.01.007. В архиве (PDF) из оригинала 14 июля 2015 г.. Получено 8 июн 2015.
- ^ Сон, Норман Х .; Занле, Кевин Дж.; Кастинг, Джеймс Ф .; и другие. (9 ноября 1989 г.). «Уничтожение экосистем из-за столкновения крупных астероидов с ранней Землей». Природа. 342 (6246): 139–142. Bibcode:1989Натура.342..139S. Дои:10.1038 / 342139a0. PMID 11536616. S2CID 1137852.
- ^ Гомес, Родни; Левисон, Хэл Ф.; Циганис, Клеоменис; Морбиделли, Алессандро (26 мая 2005 г.). «Происхождение катастрофического периода поздних тяжелых бомбардировок планет земной группы». Природа. 435 (7041): 466–469. Bibcode:2005Натура.435..466Г. Дои:10.1038 / природа03676. PMID 15917802.
- ^ Дэвис 2007, стр. 61–73
- ^ Maher, Kevin A .; Стивенсон, Дэвид Дж. (18 февраля 1988 г.). «Воздействие расстройства на зарождение жизни». Природа. 331 (6157): 612–614. Bibcode:1988Натура.331..612М. Дои:10.1038 / 331612a0. PMID 11536595. S2CID 4284492.
- ^ Чиба, Кристофер; Саган, Карл (9 января 1992 г.). «Эндогенное производство, экзогенная доставка и ударно-шоковый синтез органических молекул: инвентарь для происхождения жизни». Природа. 355 (6356): 125–132. Bibcode:1992Натура.355..125C. Дои:10.1038 / 355125a0. PMID 11538392. S2CID 4346044.
- ^ Фурукава, Ёсихиро; Секин, Тошимори; Оба, Масахиро; и другие. (Январь 2009 г.). «Образование биомолекул в результате воздействия океана на раннюю Землю». Природа Геонауки. 2 (1): 62–66. Bibcode:2009НатГе … 2 … 62F. Дои:10.1038 / NGEO383.
- ^ Дэвис 1999, п. 155
- ^ Бок и Гуд 1996
- ^ Мортильяро, Николь (1 марта 2017 г.). «Древнейшие следы жизни на Земле, найденные в Квебеке, датируются примерно 3,8 года назад». CBC News. В архиве из оригинала на 1 марта 2017 г.. Получено 2 марта 2017.
- ^ Отомо, Йоко; Какегава, Такеши; Исида, Акизуми; и другие. (Январь 2014). «Доказательства биогенного графита в метаосадочных породах Исуа раннего архея». Природа Геонауки. 7 (1): 25–28. Bibcode:2014НатГе … 7 … 25О. Дои:10.1038 / ngeo2025.
- ^ Боренштейн, Сет (13 ноября 2013 г.). «Самое древнее найденное ископаемое: познакомьтесь со своей мамой-микробом». Возбудить. Йонкерс, штат Нью-Йорк: Интерактивная сеть Mindspark. Ассошиэйтед Пресс. В архиве из оригинала 29 июня 2015 г.. Получено 2 июн 2015.
- ^ Ноффке, Нора; Кристиан, Даниэль; Уэйси, Дэвид; Хейзен, Роберт М. (16 ноября 2013 г.). «Микробиологически индуцированные осадочные структуры, фиксирующие древнюю экосистему в ок. 3.48 Формация Гио-Дрессер, Пилбара, Западная Австралия «. Астробиология. 13 (12): 1103–1124. Bibcode:2013AsBio..13.1103N. Дои:10.1089 / аст.2013.1030. ЧВК 3870916. PMID 24205812.
- ^ Уэйд, Николас (31 августа 2016 г.). «Самые старые ископаемые в мире найдены в Гренландии». Нью-Йорк Таймс. В архиве с оригинала 31 августа 2016 г.. Получено 31 августа 2016.
- ^ Дэвис 1999
- ^ Хассенкам, Т .; Андерссон, М.П .; Dalby, K.N .; Mackenzie, D.M.A .; Розинг, М. (2017). «Элементы эоархейской жизни, заключенные в минеральные включения». Природа. 548 (7665): 78–81. Bibcode:2017Натура 548 … 78H. Дои:10.1038 / природа23261. PMID 28738409. S2CID 205257931.
- ^ Перлман, Джонатан (13 ноября 2013 г.). «Найдены древнейшие признаки жизни на Земле». Дейли Телеграф. Лондон. В архиве из оригинала 16 декабря 2014 г.. Получено 15 декабря 2014.
- ^ О’Донохью, Джеймс (21 августа 2011 г.). «Самые старые надежные окаменелости показывают, что ранней жизнью был пляж». Новый ученый. 211: 13. Дои:10.1016 / S0262-4079 (11) 62064-2. В архиве с оригинала от 30 июня 2015 г.
- ^ Уэйси, Дэвид; Килберн, Мэтт Р .; Сондерс, Мартин; и другие. (Октябрь 2011 г.). «Микрофоссилии клеток, метаболизирующих серу, в скалах Западной Австралии возрастом 3,4 миллиарда лет». Природа Геонауки. 4 (10): 698–702. Bibcode:2011NatGe … 4..698Вт. Дои:10.1038 / ngeo1238.
- ^ Боренштейн, Сет (19 октября 2015 г.). «Намеки жизни на том, что считалось пустынной на ранней Земле». AP Новости. Ассошиэйтед Пресс. Получено 9 октября 2018.
- ^ Белл, Элизабет А .; Бохнике, Патрик; Харрисон, Т. Марк; и другие. (19 октября 2015 г.). «Потенциально биогенный углерод, сохранившийся в цирконе возрастом 4,1 миллиарда лет». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 112 (47): 14518–14521. Bibcode:2015ПНАС..11214518Б. Дои:10.1073 / pnas.1517557112. ЧВК 4664351. PMID 26483481. Раннее издание, опубликованное в Интернете до печати.
- ^ Вольперт, Стюарт (19 октября 2015 г.). «Жизнь на Земле, вероятно, зародилась, по крайней мере, 4,1 миллиарда лет назад — намного раньше, чем думали ученые». ULCA. В архиве из оригинала 20 октября 2015 г.. Получено 20 октября 2015.
- ^ Викрамасингхе, Чандра (2011). «Бактериальные морфологии, поддерживающие кометную панспермию: переоценка». Международный журнал астробиологии. 10 (1): 25–30. Bibcode:2011IJAsB..10 … 25 Вт. CiteSeerX 10.1.1.368.4449. Дои:10.1017 / S1473550410000157.
- ^ Рампелотто, П. Х. (2010). «Панспермия: перспективное направление исследований». В: Научная конференция по астробиологии. Abs 5224.
- ^ Чанг, Кеннет (12 сентября 2016 г.). «Видения жизни на Марсе в глубинах Земли». Нью-Йорк Таймс. В архиве из оригинала 12 сентября 2016 г.. Получено 12 сентября 2016.
- ^ Кларк, Стюарт (25 сентября 2002 г.). «Жесткий жук Земли может быть с Марса». Новый ученый. В архиве из оригинала 2 декабря 2014 г.. Получено 21 июн 2015.
- ^ Хорнек, Герда; Клаус, Дэвид М .; Манчинелли, Рокко Л. (март 2010 г.). «Космическая микробиология». Обзоры микробиологии и молекулярной биологии. 74 (1): 121–156. Bibcode:2010MMBR … 74..121H. Дои:10.1128 / MMBR.00016-09. ЧВК 2832349. PMID 20197502.
- ^ Раббоу, Элке; Хорнек, Герда; Реттберг, Петра; и другие. (Декабрь 2009 г.). «EXPOSE, установка для астробиологической экспозиции на Международной космической станции — от предложения до полета». Истоки жизни и эволюция биосфер. 39 (6): 581–598. Bibcode:2009OLEB … 39..581R. Дои:10.1007 / s11084-009-9173-6. PMID 19629743. S2CID 19749414.
- ^ Онофри, Сильвано; де ла Торре, Роза; де Вера, Жан-Пьер; и другие. (Май 2012 г.). «Выживание организмов, колонизирующих горные породы, через 1,5 года пребывания в открытом космосе». Астробиология. 12 (5): 508–516. Bibcode:2012AsBio..12..508O. Дои:10.1089 / аст.2011.0736. PMID 22680696.
- ^ Стрикленд, Эшли (26 августа 2020 г.). «Согласно новому исследованию, бактерии с Земли могут выжить в космосе и выдержать полет на Марс».. Новости CNN. Получено 26 августа 2020.
- ^ Кавагути, Юко; и другие. (26 августа 2020 г.). «Повреждение ДНК и время выживания гранул деинококковых клеток в течение 3 лет пребывания в открытом космосе». Границы микробиологии. 11: 2050. Дои:10.3389 / fmicb.2020.02050. ЧВК 7479814. PMID 32983036. S2CID 221300151.
- ^ Лоеб, Авраам (2014). «Обитаемая эпоха ранней Вселенной». Международный журнал астробиологии. 13 (4): 337–339. arXiv:1312.0613. Bibcode:2014IJAsB..13..337L. CiteSeerX 10.1.1.748.4820. Дои:10.1017 / S1473550414000196. S2CID 2777386.
- ^ Дрейфус, Клаудия (2 декабря 2014 г.). «Много обсуждаемые взгляды, которые восходят к прошлому». Нью-Йорк Таймс. Нью-Йорк. п. D2. В архиве из оригинала от 3 декабря 2014 г.. Получено 3 декабря 2014.
- ^ Циммер, Карл (12 сентября 2013 г.). «Широкая возможность возникновения жизни». Нью-Йорк Таймс. Нью-Йорк. В архиве из оригинала 8 июля 2015 г.. Получено 15 июн 2015.
- ^ Уэбб, Ричард (29 августа 2013 г.). «Изначальным бульоном жизни был сухой марсианский суп из чашки». Новый ученый. В архиве из оригинала 24 апреля 2015 г.. Получено 16 июн 2015.
- ^ Вентао Ма; Чунву Ю; Вентао Чжан; и другие. (Ноябрь 2007 г.). «Рибозимы нуклеотидсинтетазы, возможно, появились первыми в мире РНК». РНК. 13 (11): 2012–2019. Дои:10.1261 / rna.658507. ЧВК 2040096. PMID 17878321.
- ^ Шелдон 2005
- ^ Леннокс 2001, стр. 229–258
- ^ Вартанян 1973, стр. 307–312
- ^ а б Бернал 1967
- ^ Бальме, Д. (1962). «Развитие биологии у Аристотеля и Теофраста: теория спонтанного зарождения». Фронезис. 7 (1–2): 91–104. Дои:10.1163 / 156852862X00052.
- ^ Росс 1652
- ^ Добелл 1960
- ^ Бондесон 1999
- ^ Левин Р., Эверс К. «Медленная смерть спонтанного поколения (1668-1859)». Архивировано из оригинал 26 апреля 2008 г.. Получено 18 апреля 2013.
- ^ Опарин 1953 г., п. 196
- ^ Тиндаль 1905, IV, XII (1876 г.), XIII (1878 г.)
- ^ «Биогенез». Хмолпедия. Анкастер, Онтарио, Канада: WikiFoundry, Inc. В архиве из оригинала 20 мая 2014 г.. Получено 19 мая 2014.
- ^ а б Хаксли 1968
- ^ Бастиан 1871 г.
- ^ Бастиан 1871 г., п.xi – xii
- ^ Абиогенез — определение. 20 апреля 2015 г. Энциклопедия астробиологии. Дои:10.1007/978-3-642-27833-4_2-4
- ^ Бахадур, Кришна (1973). «Фотохимическое образование самоподдерживающихся коацерватов» (PDF). Труды Индийской национальной академии наук. 39B (4): 455–467. Дои:10.1016 / S0044-4057 (75) 80076-1. PMID 1242552. Архивировано из оригинал (PDF) 19 октября 2013 г.
- Бахадур, Кришна (1975). «Фотохимическое образование самоподдерживающихся коацерватов». Zentralblatt für Bakteriologie, Parasitenkunde, Infektionskrankheiten und Hygiene. 130 (3): 211–218. Дои:10.1016 / S0044-4057 (75) 80076-1. OCLC 641018092. PMID 1242552.
- ^ Кастинг 1993, п. 922
- ^ Кастинг 1993, п. 920
- ^ Бернал 1967, Происхождение жизни (А.И. Опарин, 1924), с. 199–234.
- ^ Опарин 1953 г.
- ^ Брайсон 2004, стр. 300–302
- ^ Шапиро 1987, п. 110
- ^ Бернал 1951
- ^ Мартин, Уильям Ф. (Январь 2003 г.). «О происхождении клеток: гипотеза эволюционного перехода от абиотической геохимии к хемоавтотрофным прокариотам и от прокариот к ядросодержащим клеткам». Фил. Пер. R. Soc. Лондон. А. 358 (1429): 59–83. Дои:10.1098 / rstb.2002.1183. ЧВК 1693102. PMID 12594918.
- ^ Бернал, Джон Десмонд (Сентябрь 1949 г.). «Физическая основа жизни». Труды физического общества, раздел А. 62 (9): 537–558. Bibcode:1949ППСА … 62..537Б. Дои:10.1088/0370-1298/62/9/301.
- ^ Кауфман 1995
- ^ Миллер, Стэнли Л. (15 мая 1953 г.). «Производство аминокислот в возможных условиях примитивной земли». Наука. 117 (3046): 528–529. Bibcode:1953Научный … 117..528М. Дои:10.1126 / science.117.3046.528. PMID 13056598.
- ^ Паркер, Эрик Т .; Cleaves, Henderson J .; Дворкин, Джейсон П .; и другие. (5 апреля 2011 г.). «Первоначальный синтез аминов и аминокислот в 1958 г. Миллер Х.2S-богатый искровой разряд ». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 108 (14): 5526–5531. Bibcode:2011ПНАС..108.5526П. Дои:10.1073 / pnas.1019191108. ЧВК 3078417. PMID 21422282.
- ^ Цюрих, Эт (29 ноября 2020 г.). «Раскрытие тайн первозданной атмосферы Земли 4,5 миллиарда лет назад и появление жизни». Получено 30 ноября 2020.
- ^ Ландау, Элизабет (12 октября 2016 г.). «Строительные блоки из строительных блоков жизни происходят из звездного света». НАСА. В архиве из оригинала 13 октября 2016 г.. Получено 13 октября 2016.
- ^ Гавлович, Сьюзан (6 ноября 2011 г.). «Органические« переносчики »углерода в межзвездных пылевых облаках? Недавно открытые диффузные межзвездные полосы». Science Daily. Роквилл, Мэриленд: ScienceDaily, LLC. В архиве из оригинала 11 июля 2015 г.. Получено 8 июн 2015. Пост перепечатан из материалов, предоставленных Рочестерский технологический институт.
- Geballe, Thomas R .; Наджарро, Франсиско; Фигер, Дональд Ф.; и другие. (10 ноября 2011 г.). «Инфракрасные диффузные межзвездные полосы в районе центра Галактики». Природа. 479 (7372): 200–202. arXiv:1111.0613. Bibcode:2011Натура.479..200Г. Дои:10.1038 / природа10527. PMID 22048316. S2CID 17223339.
- ^ Клайс 2001
- ^ «биологическое изобилие элементов». Энциклопедия науки. Данди, Шотландия: Дэвид Дарлинг Энтерпрайзис. В архиве из оригинала 4 февраля 2012 г.. Получено 9 октября 2008.
- ^ а б c d е Гувер, Рэйчел (21 февраля 2014 г.). «Нужно отслеживать органические наночастицы по всей Вселенной? У НАСА есть для этого приложение». Исследовательский центр Эймса. Маунтин-Вью, Калифорния: НАСА. В архиве из оригинала 6 сентября 2015 г.. Получено 22 июн 2015.
- ^ Чанг, Кеннет (18 августа 2009 г.). «Из далекой кометы — ключ к жизни». Нью-Йорк Таймс. Нью-Йорк. п. A18. В архиве из оригинала от 23 июня 2015 г.. Получено 22 июн 2015.
- ^ Гончарук, Владислав В .; Зуй, О. В. (февраль 2015 г.). «Вода и углекислый газ как главные предшественники органического вещества на Земле и в космосе». Журнал химии воды и технологий. 37 (1): 2–3. Дои:10.3103 / S1063455X15010026. S2CID 97965067.
- ^ Абу Мрад, Нинетт; Виноградов, Василиса; Дюверне, Фабрис; и другие. (2015). «Лабораторное экспериментальное моделирование: химическая эволюция органического вещества из аналогов межзвездного и кометного льда». Bulletin de la Société Royale des Sciences de Liège. 84: 21–32. Bibcode:2015BSRSL..84 … 21А. В архиве из оригинала 13 апреля 2015 г.. Получено 6 апреля 2015.
- ^ а б c Московиц, Клара (29 марта 2012 г.). «Строительные блоки жизни могли образоваться в пыли вокруг молодого солнца». Space.com. Солт-Лейк-Сити, Юта: Purch. В архиве из оригинала 14 августа 2012 г.. Получено 30 марта 2012.
- ^ «‘В комете обнаружено химическое вещество жизни «. Новости BBC. Лондон. 18 августа 2009 г. В архиве из оригинала 25 мая 2015 г.. Получено 23 июн 2015.
- ^ Томпсон, Уильям Рид; Murray, B.G .; Харе, Бишун Нараин; Саган, Карл (30 декабря 1987 г.). «Окрашивание и потемнение клатрата метана и других льдов при облучении заряженными частицами: приложения к внешней Солнечной системе». Журнал геофизических исследований. 92 (A13): 14933–14947. Bibcode:1987JGR …. 9214933T. Дои:10.1029 / JA092iA13p14933. PMID 11542127.
- ^ Старк, Энн М. (5 июня 2013 г.). «Жизнь на Земле шокирующим образом исходит из этого мира». Ливермор, Калифорния: Национальная лаборатория Лоуренса Ливермора. В архиве из оригинала 16 сентября 2015 г.. Получено 23 июн 2015.
- ^ Гольдман, Нир; Тэмблин, Исаак (20 июня 2013 г.). «Пребиотическая химия в простой ледяной смеси». Журнал физической химии А. 117 (24): 5124–5131. Bibcode:2013JPCA..117.5124G. Дои:10.1021 / jp402976n. PMID 23639050.
- ^ а б c Кэри, Бьорн (18 октября 2005 г.). «Строительные блоки жизни изобилуют в космосе»‘«. Space.com. Уотсонвилл, Калифорния: Имагинова. В архиве из оригинала 26 июня 2015 г.. Получено 23 июн 2015.
- ^ а б c Hudgins, Douglas M .; Bauschlicher, Charles W. Jr .; Алламандола, Луи Дж. (10 октября 2005 г.). «Вариации положения пика межзвездной эмиссии 6,2 мкм: индикатор N в межзвездной популяции полициклических ароматических углеводородов». Астрофизический журнал. 632 (1): 316–332. Bibcode:2005ApJ … 632..316H. CiteSeerX 10.1.1.218.8786. Дои:10.1086/432495.
- ^ а б c Des Marais, Дэвид Дж .; Allamandola, Louis J .; Сэндфорд, Скотт; и другие. (2009). «Космическое распространение химической сложности». Исследовательский центр Эймса. Маунтин-Вью, Калифорния: НАСА. Архивировано из оригинал 27 февраля 2014 г.. Получено 24 июн 2015. См. Годовой отчет исследовательского центра Эймса за 2009 г. Институт астробиологии НАСА здесь «Архивная копия». Архивировано из оригинал 1 марта 2013 г.. Получено 24 июн 2015.CS1 maint: заархивированная копия как заголовок (связь).
- ^ а б Гарсиа-Эрнандес, Доминго. А .; Манчадо, Артуро; Гарсиа-Ларио, Педро; и другие. (20 ноября 2010 г.). «Образование фуллеренов в H-содержащих планетарных туманностях». Письма в астрофизический журнал. 724 (1): L39 – L43. arXiv:1009.4357. Bibcode:2010ApJ … 724L..39G. Дои:10.1088 / 2041-8205 / 724/1 / L39. S2CID 119121764.
- ^ Витт, Адольф Н .; Vijh, Uma P .; Гордон, Карл Д. (январь 2004 г.). Обнаружение голубой флуоресценции полициклическими молекулами ароматических углеводородов в красном прямоугольнике. Заседание Американского астрономического общества 203. Атланта, Джорджия: Американское астрономическое общество. Bibcode:2003AAS … 20311017W. Архивировано из оригинал 19 декабря 2003 г.. Получено 16 января 2019.
- ^ д’Искья, Марко; Манини, Паола; Мораччи, Марко; Саладино, Рафаэле; Болл, Винсент; Тиссен, Гельмут; Эванс, Ричард А .; Пуццарини, Кристина; Бароне, Винченцо (21 августа 2019 г.). «Астрохимия и астробиология: материаловедение в стране чудес?». Международный журнал молекулярных наук. 20 (17): 4079. Дои:10.3390 / ijms20174079. ISSN 1422-0067. ЧВК 6747172. PMID 31438518.
- ^ а б «НАСА готовит ледяную органику, чтобы имитировать происхождение жизни». Space.com. Огден, Юта: Purch. 20 сентября 2012 г. В архиве с оригинала 25 июня 2015 г.. Получено 26 июн 2015.
- ^ а б Gudipati, Murthy S .; Руи Ян (1 сентября 2012 г.). «Исследование на месте радиационной обработки органических веществ в астрофизических аналогах льда — новые методы лазерной десорбции, лазерной ионизации, времяпролетные масс-спектроскопические исследования». Письма в астрофизический журнал. 756 (1): L24. Bibcode:2012ApJ … 756L..24G. Дои:10.1088 / 2041-8205 / 756/1 / L24.
- ^ «База данных ИК-спектроскопии ПАУ Эймса НАСА». НАСА. В архиве из оригинала 29 июня 2015 г.. Получено 17 июн 2015.
- ^ а б Галлори, Энцо (июнь 2011 г.). «Астрохимия и происхождение генетического материала». Рендиконти Линчеи. 22 (2): 113–118. Дои:10.1007 / s12210-011-0118-4. S2CID 96659714. «Доклад, представленный на симпозиуме« Астрохимия: молекулы в пространстве и времени »(Рим, 4–5 ноября 2010 г.), спонсором которого является Fondazione« Guido Donegani », Accademia Nazionale dei Lincei».
- ^ Мартинс, Зита (Февраль 2011 г.). «Органическая химия углеродистых метеоритов». Элементы. 7 (1): 35–40. Дои:10.2113 / gselements.7.1.35.
- ^ Мартинс, Зита; Ботта, Оливер; Фогель, Мэрилин Л .; и другие. (15 июня 2008 г.). «Внеземные азотистые основания в метеорите Мерчисон». Письма по науке о Земле и планетах. 270 (1–2): 130–136. arXiv:0806.2286. Bibcode:2008E и PSL.270..130M. Дои:10.1016 / j.epsl.2008.03.026. S2CID 14309508.
- ^ «Мы все можем быть космическими пришельцами: учеба». Сидней: Австралийская радиовещательная корпорация. Агентство Франс-Пресс. 14 июня 2008 г. В архиве из оригинала от 23 июня 2015 г.. Получено 22 июн 2015.
- ^ Каллахан, Майкл П .; Смит, Карен Э .; Кливз, Х. Джеймс, II; и другие. (23 августа 2011 г.). «Углеродистые метеориты содержат широкий спектр внеземных азотистых оснований». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 108 (34): 13995–13998. Bibcode:2011PNAS..10813995C. Дои:10.1073 / pnas.1106493108. ЧВК 3161613. PMID 21836052.
- ^ Стейгервальд, Джон (8 августа 2011 г.). «Исследователи НАСА: строительные блоки ДНК могут быть созданы в космосе». Центр космических полетов Годдарда. Гринбелт, Мэриленд: НАСА. В архиве из оригинала от 23 июня 2015 г.. Получено 23 июн 2015.
- ^ а б Чоу, Дениз (26 октября 2011 г.). «Открытие: космическая пыль содержит органическое вещество звезд». Space.com. Огден, Юта: Purch. В архиве из оригинала 14 июля 2015 г.. Получено 23 июн 2015.
- ^ Университет Гонконга (27 октября 2011 г.). «Астрономы обнаруживают, что сложные органические вещества существуют по всей Вселенной». Роквилл, Мэриленд: ScienceDaily, LLC. В архиве из оригинала от 3 июля 2015 г.
- ^ Сун Квок; Юн Чжан (3 ноября 2011 г.). «Смешанные ароматические и алифатические органические наночастицы как носители неидентифицированных характеристик инфракрасного излучения». Природа. 479 (7371): 80–83. Bibcode:2011Натура 479 … 80 тыс.. Дои:10.1038 / природа10542. PMID 22031328. S2CID 4419859.
- ^ а б Клеменс, Лара; Коэн, Джарретт (7 февраля 2005 г.). «Космический сахар — сладкая находка». Центр космических полетов Годдарда. Гринбелт, Мэриленд: НАСА. В архиве из оригинала 5 марта 2016 г.. Получено 23 июн 2015.
- ^ Тан, Кер (30 августа 2012 г.). «Сахар, найденный в космосе: признак жизни?». Новости National Geographic. Вашингтон, округ Колумбия.: Национальное географическое общество. В архиве из оригинала 14 июля 2015 г.. Получено 23 июн 2015.
- ^ «Сладко! Астрономы заметили молекулу сахара возле звезды». Возбудить. Йонкерс, штат Нью-Йорк: Интерактивная сеть Mindspark. Ассошиэйтед Пресс. 29 августа 2012 г. В архиве из оригинала 14 июля 2015 г.. Получено 23 июн 2015.
- ^ «Строительные блоки жизни найдены вокруг молодой звезды». Новости и события. Лейден, Нидерланды: Лейденский университет. 30 сентября 2012 г. В архиве из оригинала 13 декабря 2013 г.. Получено 11 декабря 2013.
- ^ Jørgensen, Jes K .; Фавр, Сесиль; Bisschop, Suzanne E .; и другие. (2012). «Обнаружение простейшего сахара, гликолевого альдегида, в протозвезде солнечного типа с помощью ALMA» (PDF). Письма в астрофизический журнал. 757 (1): L4. arXiv:1208.5498. Bibcode:2012ApJ … 757L … 4J. Дои:10.1088 / 2041-8205 / 757/1 / L4. S2CID 14205612. В архиве (PDF) из оригинала 24 сентября 2015 г.. Получено 23 июн 2015.
- ^ а б Фурукава, Ёсихиро; Чикараиси, Ёсито; Окоучи, Наохико; Ogawa, Nanako O .; Glavin, Daniel P .; Дворкин, Джейсон П .; Абэ, Чиаки; Накамура, Томоки (13 ноября 2019 г.). «Внеземная рибоза и другие сахара в примитивных метеоритах». Труды Национальной академии наук. 116 (49): 24440–24445. Bibcode:2019PNAS..11624440F. Дои:10.1073 / pnas.1907169116. ISSN 0027-8424. ЧВК 6900709. PMID 31740594.
- ^ Браун, Майкл Р. У .; Корнберг, Артур (16 ноября 2004 г.). «Неорганический полифосфат в происхождении и выживании видов». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 101 (46): 16085–16087. Bibcode:2004ПНАС..10116085Б. Дои:10.1073 / pnas.0406909101. ЧВК 528972. PMID 15520374.
- ^ Кларк, Дэвид П. (3 августа 1999 г.). «Происхождение жизни». Микробиология 425: Биохимия и физиология микроорганизмов (Лекция). Карбондейл, Иллинойс: Научный колледж; Университет Южного Иллинойса Карбондейл. Архивировано из оригинал 2 октября 2000 г.. Получено 26 июн 2015.
- ^ Пасек, Мэтью А. (22 января 2008 г.). «Переосмысление геохимии фосфора ранней Земли». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 105 (3): 853–858. Bibcode:2008ПНАС..105..853П. Дои:10.1073 / pnas.0708205105. ЧВК 2242691. PMID 18195373.
- ^ Ciesla, F.J .; Сэндфорд, С.А. (29 марта 2012 г.). «Органический синтез путем облучения и нагревания ледяных зерен в солнечной туманности». Наука. 336 (6080): 452–454. Bibcode:2012Наука … 336..452C. Дои:10.1126 / наука.1217291. HDL:2060/20120011864. PMID 22461502. S2CID 25454671.
- ^ Стейгервальд, Билл; Джонс, Нэнси; Фурукава, Ёсихиро (18 ноября 2019 г.). «Первое обнаружение сахаров в метеоритах дает ключ к разгадке происхождения жизни». НАСА. Получено 18 ноября 2019.
- ^ а б Уолш, Дж. Брюс (1995). «Часть 4: Экспериментальные исследования происхождения жизни». Истоки жизни (Конспект лекций). Тусон, Аризона: Университет Аризоны. Архивировано из оригинал 13 января 2008 г.. Получено 8 июн 2015.
- ^ а б Саладино, Рафаэле; Крестини, Клаудиа; Пино, Саманта; и другие. (Март 2012 г.). «Формамид и происхождение жизни» (PDF). Обзоры физики жизни. 9 (1): 84–104. Bibcode:2012ФЛРв … 9 … 84С. Дои:10.1016 / j.plrev.2011.12.002. HDL:2108/85168. PMID 22196896.
- ^ а б Саладино, Рафаэле; Ботта, Джорджия; Пино, Саманта; и другие. (Июль 2012 г.). «От одноуглеродного амидного формамида до РНК все стадии пребиотически возможны». Биохимия. 94 (7): 1451–1456. Дои:10.1016 / j.biochi.2012.02.018. PMID 22738728.
- ^ Марлер, Рут, изд. (3 марта 2015 г.). «НАСА Эймс воспроизводит строительные блоки жизни в лаборатории». Исследовательский центр Эймса. Моффетт Филд, Калифорния: НАСА. В архиве из оригинала 5 марта 2015 г.. Получено 5 марта 2015.
- ^ Ферус, Мартин; Несворны, Давид; Шпонер, Иржи; Кубелик, Петр; Михальчикова Регина; Шестовская, Виолетта; Šponer, Judit E .; Цивиш, Святополк (2015). «Высокоэнергетическая химия формамида: единый механизм образования азотистых оснований». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 112 (3): 657–662. Bibcode:2015ПНАС..112..657F. Дои:10.1073 / pnas.1412072111. ЧВК 4311869. PMID 25489115.
- ^ Оро, Жанна (16 сентября 1961 г.). «Механизм синтеза аденина из цианистого водорода в возможных примитивных земных условиях». Природа. 191 (4794): 1193–1194. Bibcode:1961Натура.191.1193O. Дои:10.1038 / 1911193a0. PMID 13731264. S2CID 4276712.
- ^ Базиль, Бренда; Ласкано, Антонио; Оро, Жоан (1984). «Пребиотические синтезы пуринов и пиримидинов». Успехи в космических исследованиях. 4 (12): 125–131. Bibcode:1984AdSpR … 4..125B. Дои:10.1016/0273-1177(84)90554-4. PMID 11537766.
- ^ Оргел, Лесли Э. (август 2004 г.). «Пересмотр пребиотика аденина: эвтектика и фотохимия». Истоки жизни и эволюция биосфер. 34 (4): 361–369. Bibcode:2004 ОЛЕБ … 34..361О. Дои:10.1023 / B: ORIG.0000029882.52156.c2. PMID 15279171. S2CID 4998122.
- ^ Робертсон, Майкл П .; Миллер, Стэнли Л. (29 июня 1995 г.). «Эффективный пребиотический синтез цитозина и урацила». Природа. 375 (6534): 772–774. Bibcode:1995Натура.375..772R. Дои:10.1038 / 375772a0. PMID 7596408. S2CID 4351012.
- ^ Фокс, Дуглас (февраль 2008 г.). «Разве жизнь развивалась во льду?». Обнаружить. В архиве с оригинала 30 июня 2008 г.. Получено 3 июля 2008.
- ^ Леви, Мэтью; Миллер, Стэнли Л .; Бринтон, Карен; Бада, Джеффри Л. (Июнь 2000 г.). «Пребиотический синтез аденина и аминокислот в европейских условиях». Икар. 145 (2): 609–613. Bibcode:2000Icar..145..609L. Дои:10.1006 / icar.2000.6365. PMID 11543508.
- ^ Менор-Сальван, Сезар; Руис-Бермеджо, Марта; Гусман, Марсело I .; Осуна-Эстебан, Сусана; Вейнтемильяс-Вердагер, Сабино (20 апреля 2009 г.). «Синтез пиримидинов и триазинов во льду: значение для пребиотической химии нуклеооснований». Химия: европейский журнал. 15 (17): 4411–4418. Дои:10.1002 / chem.200802656. PMID 19288488.
- ^ Рой, Дебджани; Наджафиан, Катаюн; фон Раге Шлейер, Поль (30 октября 2007 г.). «Химическая эволюция: механизм образования аденина в пребиотических условиях». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 104 (44): 17272–17277. Bibcode:2007ПНАС..10417272Р. Дои:10.1073 / pnas.0708434104. ЧВК 2077245. PMID 17951429.
- ^ а б Кливз, Х. Джеймс; Чалмерс, Джон Х .; Ласкано, Антонио; и другие. (Апрель 2008 г.). «Переоценка пребиотического органического синтеза в нейтральных планетных атмосферах». Истоки жизни и эволюция биосфер. 38 (2): 105–115. Bibcode:2008OLEB … 38..105C. Дои:10.1007 / s11084-007-9120-3. PMID 18204914. S2CID 7731172.
- ^ Чиба, Кристофер Ф. (13 мая 2005 г.). «Переосмысление ранней атмосферы Земли». Наука. 308 (5724): 962–963. Дои:10.1126 / science.1113157. PMID 15890865. S2CID 93303848.
- ^ Barton et al. 2007 г., стр. 93–95
- ^ Bada & Lazcano 2009, стр. 56–57
- ^ Bada, Джеффри Л .; Ласкано, Антонио (2 мая 2003 г.). «Суп с пребиотиками — возвращаясь к эксперименту Миллера» (PDF). Наука. 300 (5620): 745–746. Дои:10.1126 / science.1085145. PMID 12730584. S2CID 93020326. В архиве (PDF) из оригинала 4 марта 2016 г.. Получено 13 июн 2015.
- ^ Сервис, Роберт Ф. (16 марта 2015 г.). «Исследователи, возможно, решили загадку происхождения жизни». Наука (Новости). Вашингтон, округ Колумбия: Американская ассоциация по развитию науки. В архиве с оригинала 12 августа 2015 г.. Получено 26 июля 2015.
- ^ а б Patel, Bhavesh H .; Персиваль, Клаудиа; Ритсон, Дугал Дж .; Даффи, Colm D .; Сазерленд, Джон Д. (Апрель 2015 г.). «Общее происхождение РНК, белков и предшественников липидов в цианосульфидном протометаболизме». Химия природы. 7 (4): 301–307. Bibcode:2015НатЧ … 7..301П. Дои:10.1038 / nchem.2202. ЧВК 4568310. PMID 25803468.
- ^ Патель и др. 2015 г., п. 302
- ^ Оро, Жанна; Кимбалл, Обри П. (февраль 1962 г.). «Синтез пуринов в возможных примитивных земных условиях: II. Промежуточные соединения пурина из цианистого водорода». Архивы биохимии и биофизики. 96 (2): 293–313. Дои:10.1016/0003-9861(62)90412-5. PMID 14482339.
- ^ Ахуджа, Мукеш, изд. (2006). «Происхождение жизни». Наука о жизни. 1. Дели: Isha Books. п. 11. ISBN 978-81-8205-386-1. OCLC 297208106.[ненадежный источник? ]
- ^ Пол, Наташа; Джойс, Джеральд Ф. (декабрь 2004 г.). «Минимальные самовоспроизводящиеся системы». Современное мнение в области химической биологии. 8 (6): 634–639. Дои:10.1016 / j.cbpa.2004.09.005. PMID 15556408.
- ^ а б Биссетт, Эндрю Дж .; Флетчер, Стивен П. (2 декабря 2013 г.). «Механизмы автокатализа». Angewandte Chemie International Edition. 52 (49): 12800–12826. Дои:10.1002 / anie.201303822. PMID 24127341.
- ^ Кауфман 1993, гл. 7
- ^ Докинз 2004
- ^ Tjivikua, T .; Баллестер, Пабло; Ребек, Джулиус младший (Январь 1990 г.). «Самовоспроизводящаяся система». Журнал Американского химического общества. 112 (3): 1249–1250. Дои:10.1021 / ja00159a057.
- ^ Браун, Малькольм В. (30 октября 1990 г.). «Химики создают молекулу с намеком на жизнь». Нью-Йорк Таймс. Нью-Йорк. В архиве из оригинала 21 июля 2015 г.. Получено 14 июля 2015.
- ^ Джиа, Тони З .; Чандру, Кухан; Хонго, Яёи; Африн, Рехана; Усуи, Томохиро; Мьодзё, Кунихиро; Кливз, Х. Джеймс (22 июля 2019 г.). «Безмембранные микрокапли полиэстера как изначальные компартменты у истоков жизни». Труды Национальной академии наук. 116 (32): 15830–15835. Дои:10.1073 / пнас.1902336116. ЧВК 6690027. PMID 31332006.
- ^ Чандру, Кухан; Мамаджанов, Ирена; Кливз, Х. Джеймс; Джиа, Тони З. (январь 2020 г.). «Полиэфиры как модельная система для построения примитивных биологических препаратов из химии небиологических пребиотиков». Жизнь. 10 (1): 6. Дои:10.3390 / life10010006. ЧВК 7175156. PMID 31963928.
- ^ Марк, Кауфман (18 июля 2019 г.). «Астробиология НАСА». astrobiology.nasa.gov.
- ^ Гуттенберг, Николай; Дева, Натаниэль; Чандру, Кухан; Шарф, Калеб; Мамаджанов, Ирена (13 ноября 2017 г.). «Массовые измерения беспорядочного химического состава необходимы для теории происхождения жизни». Философские труды Королевского общества A: математические, физические и инженерные науки. 375 (2109): 20160347. Bibcode:2017RSPTA.37560347G. Дои:10.1098 / rsta.2016.0347. ЧВК 5686404. PMID 29133446.
- ^ Вудворд 1969, п. 287
- ^ Ланцет, Дорон (30 декабря 2014 г.). «Системная пребиология — исследования происхождения жизни». Лаборатория Ланцет. Реховот, Израиль: Отдел молекулярной генетики; Институт науки Вейцмана. В архиве из оригинала 26 июня 2015 г.. Получено 26 июн 2015.
- ^ Сегре, Даниэль; Бен-Эли, Дафна; Димер, Дэвид В .; Ланцет, Дорон (февраль 2001 г.). «Липидный мир» (PDF). Истоки жизни и эволюция биосферы. 31 (1–2): 119–145. Bibcode:2001ОЛЕБ … 31..119С. Дои:10.1023 / А: 1006746807104. PMID 11296516. S2CID 10959497. В архиве (PDF) из оригинала 26 июня 2015 г.. Получено 11 сентября 2008.
- ^ а б c Чен, Ирен А .; Вальде, Питер (июль 2010 г.). «От самоорганизующихся везикул к протоклеткам». Перспективы Колд-Спринг-Харбор в биологии. 2 (7): a002170. Дои:10.1101 / cshperspect.a002170. ЧВК 2890201. PMID 20519344.
- ^ Эйген, Манфред; Шустер, Питер (Ноябрь 1977 г.). «Гиперцикл. Принцип естественной самоорганизации. Часть A: Возникновение гиперцикла» (PDF). Naturwissenschaften. 64 (11): 541–65. Bibcode:1977NW ….. 64..541E. Дои:10.1007 / bf00450633. PMID 593400. Архивировано из оригинал (PDF) 3 марта 2016 г.. Получено 13 июн 2015.
- Эйген, Манфред; Шустер, Питер (1978). «Гиперцикл. Принцип естественной самоорганизации. Часть B: Абстрактный гиперцикл» (PDF). Naturwissenschaften. 65 (1): 7–41. Bibcode:1978NW ….. 65 …. 7E. Дои:10.1007 / bf00420631. S2CID 1812273. Архивировано из оригинал (PDF) 3 марта 2016 г.. Получено 13 июн 2015.
- Эйген, Манфред; Шустер, Питер (июль 1978). «Гиперцикл. Принцип естественной самоорганизации. Часть C: Реалистичный гиперцикл» (PDF). Naturwissenschaften. 65 (7): 341–369. Bibcode:1978NW ….. 65..341E. Дои:10.1007 / bf00439699. S2CID 13825356. Архивировано из оригинал (PDF) 16 июня 2016 г.. Получено 13 июн 2015.
- ^ Маркович, Омер; Ланцет, Дорон (лето 2012 г.). «Для эффективной эволюционируемости необходим избыточный взаимный катализ». Искусственная жизнь. 18 (3): 243–266. Дои:10.1162 / artl_a_00064. PMID 22662913. S2CID 5236043.
- ^ Тессера, Марк (2011). «Происхождение эволюции против Происхождение жизни: изменение парадигмы ». Международный журнал молекулярных наук. 12 (6): 3445–3458. Дои:10.3390 / ijms12063445. ЧВК 3131571. PMID 21747687. Спецвыпуск: «Происхождение жизни 2011»
- ^ «Изучение происхождения жизни: протоклетки». Изучение истоков жизни: виртуальная выставка. Округ Арлингтон, Вирджиния: Национальный научный фонд. В архиве из оригинала 28 февраля 2014 г.. Получено 18 марта 2014.
- ^ а б c Чен, Ирен А. (8 декабря 2006 г.). «Появление клеток в процессе зарождения жизни». Наука. 314 (5805): 1558–1559. Дои:10.1126 / science.1137541. PMID 17158315.
- ^ а б Циммер, Карл (26 июня 2004 г.). «Что было до ДНК?». Обнаружить. В архиве из оригинала 19 марта 2014 г.
- ^ Шапиро, Роберт (Июнь 2007 г.). «Более простой источник жизни». Scientific American. 296 (6): 46–53. Bibcode:2007SciAm.296f..46S. Дои:10.1038 / scientificamerican0607-46. PMID 17663224. В архиве из оригинала 14 июня 2015 г.. Получено 15 июн 2015.
- ^ Чанг 2007
- ^ Маршалл, Майкл (14 декабря 2020 г.). «Возможно, он нашел ключ к истокам жизни. Так почему же так мало о нем слышали?» — венгерский биолог Тибор Ганти — малоизвестная фигура. Теперь, спустя более десяти лет после его смерти, его идеи о том, как зародилась жизнь, наконец, идет к осуществлению «. Национальное географическое общество. Получено 15 декабря 2020.
- ^ Хьюз Берсини (2011). «Минимальная ячейка: взгляд компьютерного ученого». В Мюриэль Гарго; Пурификасьон Лопес-Гарсия; Эрве Мартин (ред.). Происхождение и эволюция жизни: астробиологическая перспектива. Издательство Кембриджского университета. С. 60–61. ISBN 9781139494595.
- ^ Ван Сегбрук С., Нове А., Ленертс Т. (2009). «Стохастическое моделирование хемотона». Artif Life. 15 (2): 213–226. CiteSeerX 10.1.1.398.8949. Дои:10.1162 / artl.2009.15.2.15203. PMID 19199383. S2CID 10634307.
- ^ Хёнигсберг ХФ (2007). «От геохимии и биохимии до пребиотической эволюции … мы обязательно входим в жидкие автоматы Ганти». Genet Mol Res. 6 (2): 358–373. PMID 17624859.
- ^ Свитек, Брайан (13 февраля 2012 г.). «Бурлящие споры о происхождении жизни». Природа. Лондон: Издательская группа Nature. Дои:10.1038 / природа.2012.10024.
- ^ z3530495 (5 мая 2020 г.). «‘Когда химия стала биологией »: в поисках истоков жизни в горячих источниках». Отдел новостей UNSW. Получено 12 октября 2020.
- ^ Дамер, Брюс; Димер, Дэвид (13 марта 2015 г.). «Парные фазы и комбинаторный отбор в колеблющихся гидротермальных бассейнах: сценарий для руководства экспериментальными подходами к происхождению клеточной жизни». Жизнь. 5 (1): 872–887. Дои:10.3390 / life5010872. ЧВК 4390883. PMID 25780958.
- ^ Гроте, Матиас (сентябрь 2011 г.). «Jeewanu, или частицы жизни‘» (PDF). Журнал биологических наук. 36 (4): 563–570. Дои:10.1007 / s12038-011-9087-0. PMID 21857103. S2CID 19551399. В архиве (PDF) из оригинала 24 сентября 2015 г.. Получено 15 июн 2015.
- ^ Гупта, В.К .; Рай, Р.К. (Август 2013). «Гистохимическая локализация РНК-подобного материала в фотохимически сформированных самоподдерживающихся абиогенных супрамолекулярных ансамблях» Дживану‘«. Международный научно-исследовательский журнал науки и техники. 1 (1): 1–4. В архиве с оригинала 28 июня 2017 г.. Получено 15 июн 2015.
- ^ Велтер, Кира (10 августа 2015 г.). «Пептидный клей мог скрепить вместе первые компоненты протоклеток». Мир химии (Новости). Лондон: Королевское химическое общество. В архиве из оригинала 5 сентября 2015 г.. Получено 29 августа 2015.
- ^ Kamat, Neha P .; Тобе, Сильвия; Хилл, Ян Т .; Шостак, Джек В. (29 июля 2015 г.). «Электростатическая локализация РНК на мембранах протоклеток с помощью катионных гидрофобных пептидов». Angewandte Chemie International Edition. 54 (40): 11735–11739. Дои:10.1002 / anie.201505742. ЧВК 4600236. PMID 26223820.
- ^ а б c Мартин, Уильям; Рассел, Майкл Дж. (29 января 2003 г.). «О происхождении клеток: гипотеза эволюционного перехода от абиотической геохимии к хемоавтотрофным прокариотам и от прокариот к ядросодержащим клеткам». Философские труды Королевского общества B. 358 (1429): 59–83, обсуждение 83–85. Дои:10.1098 / rstb.2002.1183. ЧВК 1693102. PMID 12594918.
- ^ «Письмо № 7471, Чарльз Дарвин Джозефу Далтону Хукеру, 1 февраля (1871 г.)». Дарвинский заочный проект. Получено 7 июля 2020.
- ^ Приску, Джон С. «Происхождение и эволюция жизни на замерзшей Земле». Округ Арлингтон, Вирджиния: Национальный фонд науки. В архиве из оригинала 18 декабря 2013 г.. Получено 1 марта 2014.
- ^ Маршалл, Майкл (11 ноября 2020 г.). «Предчувствие Чарльза Дарвина относительно ранней жизни, вероятно, было правильным — в нескольких нацарапанных заметках другу биолог Чарльз Дарвин предположил, как зародилась жизнь. Мало того, что это, вероятно, было правильно, его теория была на столетие раньше своего времени». Новости BBC. Получено 11 ноября 2020.
- ^ Джонстон, Ян (2 октября 2017 г.). «Впервые жизнь зародилась в« теплых прудах », почти таких же старых, как сама Земля — знаменитая идея Дарвина, подтвержденная новым научным исследованием». Независимый. В архиве из оригинала 3 октября 2017 г.. Получено 2 октября 2017.
- ^ Доктор медицины> Бразье (2012), «Секретные камеры: внутренняя история клеток и сложной жизни» (Oxford Uni Press), стр.298
- ^ Уорд, Питер и Киршвинк, Джо, op cit, p. 42
- ^ а б c d е Colín-García, M .; A. Heredia; Г. Кордеро; А. Кампруби; А. Негрон-Мендоса; Ф. Ортега-Гутьеррес; Х. Беральд; С. Рамос-Бернал (2016). «Гидротермальные источники и пребиотическая химия: обзор». Boletín de la Sociedad Geológica Mexicana. 68 (3): 599–620. Дои:10.18268 / BSGM2016v68n3a13. В архиве с оригинала 18 августа 2017 года.
- ^ Ширбер, Майкл (24 июня 2014 г.). «Гидротермальные источники могут объяснить химические прекурсоры жизни». НАСА Астробиология: Жизнь во Вселенной. НАСА. Архивировано из оригинал 29 ноября 2014 г.. Получено 19 июн 2015.
- ^ а б Переулок 2009
- ^ Игнатов, Игнат; Мосин, Олег В. (2013). «Возможные процессы происхождения жизни и живой материи с моделированием физиологических процессов бактерий». Bacillus Subtilis в тяжелой воде как модельной системе ». Журнал исследований естественных наук. 3 (9): 65–76.
- ^ Кальвин 1969
- ^ Ширбер, Майкл (1 марта 2010 г.). «Первые ископаемые в горячей воде». Журнал Astrobiology. В архиве из оригинала 14 июля 2015 г.. Получено 19 июн 2015.
- ^ Курихара, Кенсуке; Тамура, Миеко; Шохда, Кох-ичиро; и другие. (Октябрь 2011 г.). «Самовоспроизведение супрамолекулярных гигантских везикул в сочетании с амплификацией инкапсулированной ДНК». Химия природы. 3 (10): 775–781. Bibcode:2011НатЧ … 3..775К. Дои:10.1038 / nchem.1127. PMID 21941249.
- ^ Ашер, Оли (27 апреля 2015 г.). «Химический состав горячих жерл морского дна может объяснить появление жизни» (Пресс-релиз). Университетский колледж Лондона. Архивировано из оригинал 20 июня 2015 г.. Получено 19 июн 2015.
- ^ Ролдан, Альберто; Холлингсворт, Натан; Роффи, Анна; Ислам, Хусн-Убайда; и другие. (Май 2015 г.). «Биологическое преобразование CO2 с помощью катализаторов из сульфида железа в устойчивых условиях». Химические коммуникации. 51 (35): 7501–7504. Дои:10.1039 / C5CC02078F. PMID 25835242. В архиве с оригинала от 20 июня 2015 г.. Получено 19 июн 2015.
- ^ Baross, J.A .; Хоффман, С. (1985). «Подводные гидротермальные источники и связанные с ними градиентные среды как места происхождения и эволюции жизни». Истоки LifeEvol. B. 15 (4): 327–345. Bibcode:1985OrLi … 15..327B. Дои:10.1007 / bf01808177. S2CID 4613918.
- ^ Russell, M.J .; Холл, А.Дж. (1997). «Возникновение жизни из пузырьков моносульфида железа на подводном гидротермальном фронте окислительно-восстановительного потенциала и pH». Журнал геологического общества. 154 (3): 377–402. Bibcode:1997JGSoc.154..377R. Дои:10.1144 / gsjgs.154.3.0377. PMID 11541234. S2CID 24792282.
- ^ Amend, J.P .; LaRowe, D.E .; McCollom, T.M .; Шок, Э. (2013). «Энергетика органического синтеза внутри и вне клетки». Фил. Пер. R. Soc. Лондон. B. 368 (1622): 20120255. Дои:10.1098 / rstb.2012.0255. ЧВК 3685458. PMID 23754809.
- ^ Shock, E.L .; Бойд, Э. (2015). «Геомикробиология и микробиологическая геохимия: основы геобиохимии». Элементы. 11: 389–394. Дои:10.2113 / gselements.11.6.395.
- ^ Martin, W .; Рассел, М.Дж. (2007). «О происхождении биохимии щелочного гидротермального источника». Фил. Пер. R. Soc. Лондон. B. 362 (1486): 1887–1925. Дои:10.1098 / rstb.2006.1881. ЧВК 2442388. PMID 17255002.
- ^ Природа, Том 535, 28 июля 2016. с.468.
- ^ Чандру, Кухан; Гуттенберг, Николай; Гири, Чайтанья; Хонго, Яёи; Бутч, Кристофер; Мамаджанов, Ирена; Кливз, Х. Джеймс (31 мая 2018 г.). «Простой пребиотический синтез разнообразных динамических комбинаторных полиэфирных библиотек». Коммуникационная химия. 1 (1). Дои:10.1038 / с42004-018-0031-1.
- ^ Форсайт, Джей Джи; Юй Шэн-Шэн; Мамаджанов, Ирена; Гровер, Марта А; Кришнамурти, Раманараянан; Фернандес, Факундо М; Худ, Николай V (17 августа 2015 г.). «Эфир-опосредованное образование амидных связей, управляемое влажно-сухими циклами: возможный путь к полипептидам на пребиотической Земле». Angewandte Chemie International Edition на английском языке. 54 (34): 9871–9875. Дои:10.1002 / anie.201503792. ЧВК 4678426. PMID 26201989.
- ^ Мулкиджанян, Армид; Бычков, Андрей; Диброва, Дарья; Гальперин, Михаил; Кунин, Евгений (3 апреля 2012 г.). «Происхождение первых ячеек на земных бескислородных геотермальных полях». PNAS. 109 (14): E821 – E830. Bibcode:2012PNAS..109E.821M. Дои:10.1073 / pnas.1117774109. ЧВК 3325685. PMID 22331915.
- ^ Хоффманн, Джеффри Уильям (24 декабря 2016 г.). «Сетевая теория происхождения жизни». bioRxiv 10.1101/096701.
- ^ Дартнелл, Льюис (12 января 2008 г.). «Жизнь началась на радиоактивном пляже?». Новый ученый (2638): 8. В архиве из оригинала 27 июня 2015 г.. Получено 26 июн 2015.
- ^ Адам, Захари (2007). «Актиниды и истоки жизни». Астробиология. 7 (6): 852–872. Bibcode:2007AsBio … 7..852A. Дои:10.1089 / аст.2006.0066. PMID 18163867.
- ^ Парнелл, Джон (декабрь 2004 г.). «Минеральная радиоактивность песков как механизм фиксации органического углерода на ранней Земле». Истоки жизни и эволюция биосфер. 34 (6): 533–547. Bibcode:2004 ОЛЕБ … 34..533П. CiteSeerX 10.1.1.456.8955. Дои:10.1023 / B: ORIG.0000043132.23966.a1. PMID 15570707. S2CID 6067448.
- ^ Дэвис, Пол (Декабрь 2007 г.). «Есть ли среди нас инопланетяне?» (PDF). Scientific American. 297 (6): 62–69. Bibcode:2007SciAm.297f..62D. Дои:10.1038 / scientificamerican1207-62. В архиве (PDF) из оригинала 4 марта 2016 г.. Получено 16 июля 2015.
… если в земных условиях жизнь действительно возникает легко, то, возможно, она много раз формировалась на нашей родной планете. Чтобы исследовать эту возможность, пустыни, озера и другие экстремальные или изолированные среды были исследованы на предмет наличия «чужеродных» форм жизни — организмов, которые фундаментально отличались бы от известных организмов, потому что они возникли независимо.
- ^ Хартман, Хайман (1998). «Фотосинтез и происхождение жизни». Истоки жизни и эволюция биосфер. 28 (4–6): 515–521. Bibcode:1998OLEB … 28..515H. Дои:10.1023 / А: 1006548904157. PMID 11536891. S2CID 2464.
- ^ а б c Сентилингам, Мира (25 апреля 2014 г.). «Метаболизм мог начаться в ранних океанах еще до зарождения жизни» (Пресс-релиз). Wellcome Trust. EurekAlert!. В архиве из оригинала 17 июня 2015 г.. Получено 16 июн 2015.
- ^ Перри, Кэролайн (7 февраля 2011 г.). «Пузыри, покрытые глиной, возможно, сформировали первые протоклетки» (Пресс-релиз). Кембридж, Массачусетс: Гарвардский университет. EurekAlert !. В архиве из оригинала 14 июля 2015 г.. Получено 20 июн 2015.
- ^ Докинз 1996, стр. 148–161
- ^ Вэньхуа Хуанг; Феррис, Джеймс П. (12 июля 2006 г.). «Одностадийный региоселективный синтез до 50-меров олигомеров РНК с помощью монтмориллонитового катализа». Журнал Американского химического общества. 128 (27): 8914–8919. Дои:10.1021 / ja061782k. PMID 16819887.
- ^ Мур, Кэролайн (16 июля 2007 г.). «Кристаллы как гены?». Основные моменты в химической науке. В архиве из оригинала 14 июля 2015 г.. Получено 21 июн 2015.
- Буллард, Тереза; Фройденталь, Джон; Авагян, Серин; и другие. (2007). «Проверка гипотезы Кэрнса-Смита« кристаллы как гены »». Фарадеевские дискуссии. 136: 231–245. Bibcode:2007FaDi..136..231B. Дои:10.1039 / b616612c. PMID 17955812.
- ^ Юэ-Чинг Хо, Юджин (июль – сентябрь 1990 г.). «Эволюционная эпистемология и последний интеллектуальный интерес сэра Карла Поппера: отчет из первых рук». Интеллектус. 15: 1–3. OCLC 26878740. В архиве из оригинала 11 марта 2012 г.. Получено 13 августа 2012.
- ^ Уэйд, Николас (22 апреля 1997 г.). «Любительские встряски идеи о рецепте жизни». Нью-Йорк Таймс. Нью-Йорк. В архиве из оригинала 17 июня 2015 г.. Получено 16 июн 2015.
- ^ Поппер, Карл Р. (29 марта 1990 г.). «Пирит и происхождение жизни». Природа. 344 (6265): 387. Bibcode:1990Натура.344..387П. Дои:10.1038 / 344387a0. S2CID 4322774.
- ^ Хубер, Клаудиа; Wächtershäuser, Günter (31 июля 1998 г.). «Пептиды путем активации аминокислот с CO на (Ni, Fe) S поверхности: значение для происхождения жизни». Наука. 281 (5377): 670–672. Bibcode:1998Sci … 281..670H. Дои:10.1126 / science.281.5377.670. PMID 9685253.
- ^ Мюссер, Джордж (23 сентября 2011 г.). «Как жизнь возникла на Земле и как сингулярность может ее разрушить». Наблюдения (Блог). В архиве из оригинала 17 июня 2015 г.. Получено 17 июн 2015.
- ^ Кэрролл, Шон (10 марта 2010 г.). «Свободная энергия и смысл жизни». Космическое отклонение (Блог). Обнаружить. В архиве из оригинала 14 июля 2015 г.. Получено 17 июн 2015.
- ^ Вулховер, Натали (22 января 2014 г.). «Новая физическая теория жизни». Журнал Quanta. В архиве из оригинала 13 июня 2015 г.. Получено 17 июн 2015.
- ^ Англия, Джереми Л. (28 сентября 2013 г.). «Статистическая физика самовоспроизведения» (PDF). Журнал химической физики. 139 (12): 121923. arXiv:1209.1179. Bibcode:2013ЖЧФ.139л1923Э. Дои:10.1063/1.4818538. HDL:1721.1/90392. PMID 24089735. S2CID 478964. В архиве (PDF) из оригинала 4 июня 2015 г.. Получено 18 июн 2015.
- ^ Оргел, Лесли Э. (7 ноября 2000 г.). «Самоорганизующиеся биохимические циклы». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 97 (23): 12503–12507. Bibcode:2000PNAS … 9712503O. Дои:10.1073 / pnas.220406697. ЧВК 18793. PMID 11058157.
- ^ Чандру, Кухан; Гилберт, Алексис; Бутч, Кристофер; Аоно, Масаси; Кливз, Хендерсон Джеймс II (21 июля 2016 г.). «Абиотическая химия производных тиолированного ацетата и происхождение жизни». Научные отчеты. 6 (29883): 29883. Bibcode:2016НатСР … 629883C. Дои:10.1038 / srep29883. ЧВК 4956751. PMID 27443234.
- ^ Валле, Янник; Шалаил, Ибрагим; Ли, Кью-Дунг; Рао, К. В. Рагхавендра; Паэпе, Гаэль Де; Меркер, Катарина; Милет, Анна (8 ноября 2017 г.). «В самом начале жизни на Земле: мировая гипотеза о богатых тиолами пептидах (TRP)». Международный журнал биологии развития. 61 (8–9): 471–478. Дои:10.1387 / ijdb.170028yv. PMID 29139533.
- ^ а б Мулкиджанян, Армен Ю. (24 августа 2009 г.). «О происхождении жизни в цинковом мире: 1. Фотосинтезирующие пористые сооружения, построенные из гидротермально осажденного сульфида цинка как колыбели жизни на Земле». Биология Директ. 4: 26. Дои:10.1186/1745-6150-4-26. ЧВК 3152778. PMID 19703272.
- ^ Wächtershäuser, Günter (декабрь 1988 г.). «До ферментов и шаблонов: теория поверхностного метаболизма». Микробиологические обзоры. 52 (4): 452–484. Дои:10.1128 / MMBR.52.4.452-484.1988. ЧВК 373159. PMID 3070320.
- ^ Мулкиджанян, Армен Ю .; Гальперин, Михаил Юрьевич (24 августа 2009 г.). «О происхождении жизни в цинковом мире. 2. Подтверждение гипотезы о фотосинтезирующих зданиях из сульфида цинка как колыбелях жизни на Земле». Биология Директ. 4: 27. Дои:10.1186/1745-6150-4-27. ЧВК 2749021. PMID 19703275.
- ^ Макаллум, А.Б. (1 апреля 1926 г.). «Палеохимия жидкостей и тканей организма». Физиологические обзоры. 6 (2): 316–357. Дои:10.1152 / Physrev.1926.6.2.316.
- ^ Мулкиджанян, Армен Ю .; Бычков, Андрей Юрьевич; Диброва, Дарья В .; и другие. (3 апреля 2012 г.). «Происхождение первых ячеек на земных бескислородных геотермальных полях». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 109 (14): E821 – E830. Bibcode:2012PNAS..109E.821M. Дои:10.1073 / pnas.1117774109. ЧВК 3325685. PMID 22331915.
- ^ Более глубокую интегративную версию этой гипотезы см., В частности, Ланкенау 2011, pp. 225–286, связывая концепцию «двух миров РНК» и другие подробные аспекты; и Давидович, Чен; Белоусов, Мэтью; Башан, Анат; Йонат, Ада (Сентябрь 2009 г.). «Эволюционирующая рибосома: от образования некодируемых пептидных связей до сложных механизмов трансляции». Исследования в области микробиологии. 160 (7): 487–492. Дои:10.1016 / j.resmic.2009.07.004. PMID 19619641.
- ^ Старр, Мишель (3 октября 2020 г.). «Новое химическое« древо происхождения жизни »раскрывает нашу возможную молекулярную эволюцию». ScienceAlert. Получено 3 октября 2020.
- ^ Волос, Агнешка; и другие. (25 сентября 2020 г.). «Синтетическая связь, возникновение и самовосстановление в сети пребиотической химии». Наука. 369 (6511). Дои:10.1126 / science.aaw1955 (неактивно 11 ноября 2020 г.). PMID 32973002. Получено 3 октября 2020.CS1 maint: DOI неактивен по состоянию на ноябрь 2020 г. (связь)
- ^ Эйген и Шустер 1979
- ^ Михаэлиан, К (2009). «Термодинамическое происхождение жизни». Динамика системы Земли. 0907 (2011): 37–51. arXiv:0907.0042. Bibcode:2011ESD ….. 2 … 37M. Дои:10.5194 / esd-2-37-2011. S2CID 14574109.
- ^ Саган, К. (1973) Давление ультрафиолетового отбора на самые ранние организмы, J. Theor. Биол., 39, 195–200.
- ^ Михаэлиан, К; Симеонов, А (2015). «Основные молекулы жизни — это пигменты, которые возникли и совместно эволюционировали в ответ на термодинамический императив рассеяния преобладающего солнечного спектра». Биогеонауки. 12 (16): 4913–4937. arXiv:1405.4059v2. Bibcode:2015BGeo … 12,49 13M. Дои:10.5194 / bg-12-4913-2015.
- ^ Михаэлиан, К (2013). «Нелинейная необратимая термодинамическая перспектива распространения органических пигментов и биологической эволюции». Journal of Physics: Серия конференций. 475 (1): 012010. arXiv:1307.5924. Bibcode:2013JPhCS.475a2010M. Дои:10.1088/1742-6596/475/1/012010. S2CID 118564759.
- ^ Doglioni, C .; Pignatti, J .; Коулман, М. (2016). «Почему жизнь возникла на поверхности Земли в кембрии?» (PDF). Границы геонаук. 7 (6): 865–873. Дои:10.1016 / j.gsf.2016.02.001.
- ^ Михаэлиан, Каро; Сантильян, Норберто (июнь 2019 г.). «UVC фотон-индуцированная денатурация ДНК: возможный диссипативный путь к бесферментной репликации архея». Гелион. 5 (6): e01902. Дои:10.1016 / j.heliyon.2019.e01902. ЧВК 6584779. PMID 31249892.
- ^ Михаэлиан, Каро (июнь 2018 г.). «Гомохиральность через индуцированную фотонами денатурацию РНК / ДНК в происхождении жизни». Жизнь. 8 (2): 21. Дои:10.3390 / life8020021. ЧВК 6027432. PMID 29882802.
- ^ Мори, CP (2009). «Саморазвивающиеся полипептидные структуры бета-листа как пребиотические информационные сущности: амилоидный мир». Истоки жизни и эволюция биосфер. 39 (2): 141–150. Дои:10.1007 / s11084-009-9165-6. PMID 19301141. S2CID 20073536.
- ^ Мори, CP (2015). «Происхождение жизни. Первичная генетика: передача информации в мире пре-РНК на основе самовоспроизводящихся амилоидных конформеров бета-листов». Журнал теоретической биологии. 382: 292–297. Дои:10.1016 / j.jtbi.2015.07.008. PMID 26196585.
- ^ Нанда, Дж; Рубинов, Б; Ивницкий, Д; Мукерджи, Р. Штельман, Э; Motro, Y; Миллер, Y; Вагнер, Н; Коэн-Лурия, Р. Ашкенасы, Г. (2017). «Появление нативных пептидных последовательностей в пребиотических сетях репликации». Nature Communications. 8 (1): 343. Bibcode:2017НатКо … 8..434N. Дои:10.1038 / s41467-017-00463-1. ЧВК 5585222. PMID 28874657.
- ^ Маршрут, СК; Фридманн, депутат; Riek, R; Гринвальд, Дж (2018). «Шаблонно-направленный синтез пребиотиков на основе амилоидов». Nature Communications. 9 (1): 234–242. Дои:10.1038 / s41467-017-02742-3. ЧВК 5770463. PMID 29339755.
- ^ Блюм, Х.Ф. (1957). О происхождении самовоспроизводящихся систем. В ритмических и синтетических процессах роста, под ред. Рудник, Д., стр. 155–170. Princeton University Press, Принстон, Нью-Джерси.
- ^ а б Токарный станок, Ричард (2004). «Быстрые приливно-отливные циклы и происхождение жизни». Икар. 168 (1): 18–22. Bibcode:2004Icar..168 … 18л. Дои:10.1016 / j.icarus.2003.10.018.
- ^ Токарный станок, Ричард (2005). «Приливная цепная реакция и происхождение реплицирующихся биополимеров». Международный журнал астробиологии. 4 (1): 19–31. Bibcode:2005IJAsB … 4 … 19л. Дои:10.1017 / S1473550405002314.
- ^ Varga, P .; Рыбицкий, К .; Денис, К. (2006). «Комментарий к статье Ричарда Лэта» Быстрые приливные циклы и происхождение жизни «. Икар. 180 (1): 274–276. Bibcode:2006Icar..180..274V. Дои:10.1016 / j.icarus.2005.04.022.
- ^ Токарный, Р. (2006). «Ранние приливы: ответ Варге и др.». Икар. 180 (1): 277–280. Bibcode:2006Icar..180..277L. Дои:10.1016 / j.icarus.2005.08.019.
- ^ Фламент, Николас; Кольтис, Николас; Рей, Патрис Ф. (2008). «Случай позднеархейского континентального возникновения на основе моделей термической эволюции и гипсометрии». Письма по науке о Земле и планетах. 275 (3–4): 326–336. Bibcode:2008E и PSL.275..326F. Дои:10.1016 / j.epsl.2008.08.029.
- ^ Мюллер, Энтони В. Дж. (7 августа 1985 г.). «Термосинтез биомембранами: получение энергии от циклических изменений температуры». Журнал теоретической биологии. 115 (3): 429–453. Дои:10.1016 / S0022-5193 (85) 80202-2. PMID 3162066.
- ^ Мюллер, Энтони В. Дж. (1995). «Были ли первые организмы тепловыми двигателями? Новая модель биогенеза и ранней эволюции преобразования биологической энергии». Прогресс в биофизике и молекулярной биологии. 63 (2): 193–231. Дои:10.1016/0079-6107(95)00004-7. PMID 7542789.
- ^ Muller, Anthonie W. J .; Шульце-Макух, Дирк (1 апреля 2006 г.). «Сорбционные тепловые машины: простые неодушевленные генераторы отрицательной энтропии». Physica A: Статистическая механика и ее приложения. 362 (2): 369–381. arXiv:физика / 0507173. Bibcode:2006PhyA..362..369M. Дои:10.1016 / j.physa.2005.12.003. S2CID 96186464.
- ^ Оргель 1987, стр. 9–16
- ^ Оргел, Лесли Э. (17 ноября 2000 г.). «Более простая нуклеиновая кислота». Наука. 290 (5495): 1306–1307. Дои:10.1126 / science.290.5495.1306. PMID 11185405. S2CID 83662769.
- ^ Нельсон, Кевин Э .; Леви, Мэтью; Миллер, Стэнли Л. (11 апреля 2000 г.). «Пептидные нуклеиновые кислоты, а не РНК, возможно, были первой генетической молекулой». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 97 (8): 3868–3871. Bibcode:2000PNAS … 97.3868N. Дои:10.1073 / pnas.97.8.3868. ЧВК 18108. PMID 10760258.
- ^ Ларральде, Роза; Робертсон, Майкл П .; Миллер, Стэнли Л. (29 августа 1995 г.). «Скорость разложения рибозы и других сахаров: последствия для химической эволюции». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 92 (18): 8158–8160. Bibcode:1995PNAS … 92.8158L. Дои:10.1073 / пнас.92.18.8158. ЧВК 41115. PMID 7667262.
- ^ Линдаль, Томас (22 апреля 1993 г.). «Неустойчивость и распад первичной структуры ДНК». Природа. 362 (6422): 709–715. Bibcode:1993Натура.362..709L. Дои:10.1038 / 362709a0. PMID 8469282. S2CID 4283694.
- ^ Анастаси, Кэрол; Кроу, Майкл А .; Паунер, Мэтью У .; Сазерленд, Джон Д. (18 сентября 2006 г.). «Прямая сборка предшественников нуклеозидов из двух- и трехуглеродных единиц». Angewandte Chemie International Edition. 45 (37): 6176–6179. Дои:10.1002 / anie.200601267. PMID 16917794.
- ^ Паунер, Мэтью У .; Сазерленд, Джон Д. (13 октября 2008 г.). «Потенциально пребиотический синтез пиримидин-β-D-рибонуклеотидов путем фотоаномеризации / гидролиза α-D-цитидин-2′-фосфата». ChemBioChem. 9 (15): 2386–2387. Дои:10.1002 / cbic.200800391. PMID 18798212. S2CID 5704391.
- ^ Паунер, Мэтью У .; Герланд, Беатрис; Сазерленд, Джон Д. (14 мая 2009 г.). «Синтез активированных пиримидин рибонуклеотидов в пребиотически вероятных условиях». Природа. 459 (7244): 239–242. Bibcode:2009Натура.459..239P. Дои:10.1038 / природа08013. PMID 19444213. S2CID 4412117.
- ^ Палашек, Стан (23 мая 2013 г.). «Репликация первичной РНК и применение в технологии ПЦР». arXiv:1305.5581v1 [q-bio.BM ].
- ^ Кунин, Евгений В.; Сенкевич, Татьяна Г .; Доля, Валериан В. (19 сентября 2006 г.). «Древний вирусный мир и эволюция клеток». Биология Директ. 1: 29. Дои:10.1186/1745-6150-1-29. ЧВК 1594570. PMID 16984643.
- ^ Власов, Александр В .; Казаков, Сергей А .; Джонстон, Брайан Х .; и другие. (Август 2005 г.). «Мир РНК на льду: новый сценарий появления информации о РНК». Журнал молекулярной эволюции. 61 (2): 264–273. Bibcode:2005JMolE..61..264V. Дои:10.1007 / s00239-004-0362-7. PMID 16044244. S2CID 21096886.
- ^ Нусинов, Марк Д .; Отрощенко, Владимир А .; Сантоли, Сальваторе (1997). «Возникновение неклеточной фазы жизни на мелкозернистых глинистых частицах реголита ранней Земли». Биосистемы. 42 (2–3): 111–118. Дои:10.1016 / S0303-2647 (96) 01699-1. PMID 9184757.
- ^ а б c Йетс, Диана (25 сентября 2015 г.). «Исследование добавляет доказательства того, что вирусы живы» (Пресс-релиз). Шампейн, Иллинойс: Университет Иллинойса в Урбане-Шампейн. В архиве из оригинала 19 ноября 2015 г.. Получено 20 октября 2015.
- ^ Янич, Александар (2018). «Необходимость включения методов обнаружения вирусов в будущие миссии на Марс». Астробиология. 18 (12): 1611–1614. Bibcode:2018AsBio..18.1611J. Дои:10.1089 / аст.2018.1851.
- ^ Кацуракис, А (2013). «Палеовирология: вывод эволюции вируса на основании данных о последовательности генома хозяина». Философские труды Королевского общества B: биологические науки. 368 (1626): 20120493. Дои:10.1098/rstb.2012.0493. ЧВК 3758182. PMID 23938747.
- ^ Arshan, Nasir; Caetano-Anollés, Gustavo (25 September 2015). «Исследование происхождения и эволюции вирусов на основе филогеномных данных». Достижения науки. 1 (8): e1500527. Bibcode:2015SciA …. 1E0527N. Дои:10.1126 / sciadv.1500527. ЧВК 4643759. PMID 26601271.
- ^ Насир, Аршан; Naeem, Aisha; Jawad Khan, Muhammad; и другие. (Декабрь 2011 г.). «Annotation of Protein Domains Reveals Remarkable Conservation in the Functional Make up of Proteomes Across Superkingdoms». Гены. 2 (4): 869–911. Дои:10.3390/genes2040869. ЧВК 3927607. PMID 24710297.
- ^ Jalasvuori M, Mattila S, Hoikkala V (2015). «Chasing the Origin of Viruses: Capsid-Forming Genes as a Life-Saving Preadaptation within a Community of Early Replicators». PLOS ONE. 10 (5): e0126094. Bibcode:2015PLoSO..1026094J. Дои:10.1371/journal.pone.0126094. ЧВК 4425637. PMID 25955384.
- ^ а б c Krupovic M, Dolja VV, Koonin EV (July 2019). «Origin of viruses: primordial replicators recruiting capsids from hosts» (PDF). Обзоры природы. Микробиология. 17 (7): 449–458. Дои:10.1038/s41579-019-0205-6. PMID 31142823. S2CID 169035711.
- ^ Orgel, Leslie E. (October 1994). «The origin of life on Earth». Scientific American. 271 (4): 76–83. Bibcode:1994SciAm.271d..76O. Дои:10.1038/scientificamerican1094-76. PMID 7524147.
- ^ Camprubí, E.; de Leeuw, J.W.; House, C.H .; Раулин, Ф .; Russell, M.J.; Spang, A .; Tirumalai, M.R.; Westall, F. (12 December 2019). «Emergence of Life». Космические науки. Rev. 215 (56): 56. Bibcode:2019SSRv..215…56C. Дои:10.1007/s11214-019-0624-8.
- ^ Johnston, Wendy K.; Unrau, Peter J.; Lawrence, Michael S.; и другие. (18 мая 2001 г.). «RNA-Catalyzed RNA Polymerization: Accurate and General RNA-Templated Primer Extension». Наука. 292 (5520): 1319–1325. Bibcode:2001Sci … 292.1319J. CiteSeerX 10.1.1.70.5439. Дои:10.1126 / science.1060786. PMID 11358999. S2CID 14174984.
- ^ Szostak, Jack W. (5 февраля 2015 г.). «The Origins of Function in Biological Nucleic Acids, Proteins, and Membranes». Chevy Chase (CDP), MD: Медицинский институт Говарда Хьюза. В архиве из оригинала 14 июля 2015 г.. Получено 16 июн 2015.
- ^ Lincoln, Tracey A.; Joyce, Gerald F. (27 February 2009). «Self-Sustained Replication of an RNA Enzyme». Наука. 323 (5918): 1229–1232. Bibcode:2009Sci…323.1229L. Дои:10.1126/science.1167856. ЧВК 2652413. PMID 19131595.
- ^ а б Joyce, Gerald F. (2009). «Evolution in an RNA world». Перспективы Колд-Спринг-Харбор в биологии. 74 (Evolution: The Molecular Landscape): 17–23. Дои:10.1101/sqb.2009.74.004. ЧВК 2891321. PMID 19667013.
- ^ а б Бернштейн, Харрис; Byerly, Henry C.; Hopf, Frederick A.; и другие. (Июнь 1983 г.). «The Darwinian Dynamic». Ежеквартальный обзор биологии. 58 (2): 185–207. Дои:10.1086/413216. JSTOR 2828805. S2CID 83956410.
- ^ а б Michod 1999
- ^ McCollom, Thomas; Mayhew, Lisa; Scott, Jim (7 October 2014). «NASA awards CU-Boulder-led team $7 million to study origins, evolution of life in universe» (Пресс-релиз). Боулдер, Колорадо: Университет Колорадо в Боулдере. Архивировано из оригинал 31 июля 2015 г.. Получено 8 июн 2015.
- ^ Oehlenschläger, Франк; Эйген, Манфред (Декабрь 1997 г.). «30 лет спустя — новый подход к эволюционным исследованиям in vitro Сола Шпигельмана и Лесли Оргеля, посвященный Лесли Оргелю по случаю его 70-летия». Истоки жизни и эволюция биосфер. 27 (5–6): 437–457. Bibcode:1997OLEB … 27..437O. Дои:10.1023 / А: 1006501326129. PMID 9394469. S2CID 26717033.
- ^ Gibson, Daniel G.; Glass, John I.; Lartigue, Carole; и другие. (2 июля 2010 г.). «Creation of a Bacterial Cell Controlled by a Chemically Synthesized Genome». Наука. 329 (5987): 52–56. Bibcode:2010Sci…329…52G. CiteSeerX 10.1.1.167.1455. Дои:10.1126/science.1190719. PMID 20488990. S2CID 7320517.
- ^ Swaby, Rachel (20 May 2010). «Scientists Create First Self-Replicating Synthetic Life». Проводной. Нью-Йорк. В архиве из оригинала 17 июня 2015 г.. Получено 8 июн 2015.
- ^ Coughlan, Andy (2016) «Smallest ever genome comes to life: Humans built it but we don’t know what a third of its genes actually do» (New Scientist 2 April 2016 No 3067)p.6
- ^ а б Балч, Эрика (4 октября 2018 г.). «Новаторская лаборатория готова раскрыть тайну происхождения жизни на Земле и за ее пределами». Университет Макмастера. Получено 4 октября 2018.
- ^ Персонал (4 октября 2018 г.). «Новаторская лаборатория готова раскрыть тайну происхождения жизни». EurekAlert!. Получено 14 октября 2018.
- ^ Персонал (2018). «Симулятор планеты». IntraVisionGroup.com. Получено 14 октября 2018.
- ^ Андерсон, Пол Скотт (14 октября 2018 г.). «New technology may help solve mystery of life’s origins – How did life on Earth begin? A new technology, called Planet Simulator, might finally help solve the mystery». EarthSky. Получено 14 октября 2018.
Источники
- Altermann, Wladyslaw (2009). «Introduction: A Roadmap to Fata Morgana?». В Зекбахе, Джозеф; Уолш, Мод (ред.). От окаменелостей до астробиологии: летописи жизни на Земле и поиск внеземных биосигнатур. Клеточное происхождение, жизнь в экстремальных средах обитания и астробиология. 12. Dordrecht, the Netherlands; Лондон: Springer Science + Business Media. ISBN 978-1-4020-8836-0. LCCN 2008933212.
- Bada, Jeffrey L.; Lazcano, Antonio (2009). «The Origin of Life». В Русе, Майкл; Travis, Joseph (ред.). Эволюция: первые четыре миллиарда лет. Предисловие Эдвард О. Уилсон. Кембридж: Belknap Press of Harvard University Press. ISBN 978-0-674-03175-3. LCCN 2008030270. OCLC 225874308.
- Barton, Nicholas H.; Бриггс, Дерек Э.; Eisen, Jonathan A.; и другие. (2007). Эволюция. Колд-Спринг-Харбор, Нью-Йорк: Лаборатория Колд-Спринг-Харбор. ISBN 978-0-87969-684-9. LCCN 2007010767. OCLC 86090399.
- Bastian, H. Charlton (1871). The Modes of Origin of Lowest Organisms. Лондон; Нью-Йорк: Макмиллан и компания. LCCN 11004276. OCLC 42959303. Получено 6 июн 2015.
- Bernal, J.D. (1951). The Physical Basis of Life. Лондон: Рутледж & Kegan Paul. LCCN 51005794.
- Bernal, J.D. (1960). «The Problem of Stages in Biopoesis». В Florkin, M. (ред.). Aspects of the Origin of Life. International Series of Monographs on Pure and Applied Biology. Оксфорд, Великобритания; Нью-Йорк: Pergamon Press. ISBN 978-1-4831-3587-8. LCCN 60013823.
- Bernal, J.D. (1967) [Reprinted work by А.И. Опарин originally published 1924; Москва: The Moscow Worker ]. Происхождение жизни. The Weidenfeld and Nicolson Natural History. Translation of Oparin by Ann Synge. Лондон: Вайденфельд и Николсон. LCCN 67098482.
- Bock, Gregory R.; Goode, Jamie A., eds. (1996). Evolution of Hydrothermal Ecosystems on Earth (and Mars?). Ciba Foundation Symposium. 202. Чичестер, Великобритания; Нью-Йорк: Джон Уайли и сыновья. ISBN 978-0-471-96509-1. LCCN 96031351.
- Бондесон, Янв (1999). Русалка Фиджи и другие очерки естественной и неестественной истории. Итака, штат Нью-Йорк: Издательство Корнельского университета. ISBN 978-0-8014-3609-3. LCCN 98038295.
- Брайсон, Билл (2004). Краткая история почти всего. Лондон: Черный лебедь. ISBN 978-0-552-99704-1. OCLC 55589795.
- Кальвин, Мелвин (1969). Chemical Evolution: Molecular Evolution Towards the Origin of Living Systems on the Earth and Elsewhere. Оксфорд, Великобритания: Clarendon Press. ISBN 978-0-19-855342-7. LCCN 70415289. OCLC 25220.
- Cardwell, Donald S. L. (1971). From Watt to Clausius: The Rise of Thermodynamics in the Early Industrial Age. Издательство Корнельского университета.
- Chaichian, Masud; Rojas, Hugo Perez; Tureanu, Anca (2014). «Physics and Life». Basic Concepts in Physics: From the Cosmos to Quarks. Конспект лекций по физике. Берлин; Heidelberg: Springer Berlin Heidelberg. Дои:10.1007/978-3-642-19598-3_12. ISBN 978-3-642-19597-6. LCCN 2013950482. OCLC 900189038.
- Chang, Thomas Ming Swi (2007). Artificial Cells: Biotechnology, Nanomedicine, Regenerative Medicine, Blood Substitutes, Bioencapsulation, and Cell/Stem Cell Therapy. Regenerative Medicine, Artificial Cells and Nanomedicine. 1. Hackensack, NJ: Всемирный научный. ISBN 978-981-270-576-1. LCCN 2007013738. OCLC 173522612.
- Dalrymple, G. Brent (2001). «Возраст Земли в двадцатом веке: проблема (в основном) решена». In Lewis, C.L.E.; Knell, S.J. (ред.). The Age of the Earth: from 4004 BC to AD 2002. Лондонское геологическое общество, специальные публикации. Специальное издание Геологического общества. 190. Лондон: Геологическое общество Лондона. С. 205–221. Bibcode:2001ГСЛСП.190..205Д. Дои:10.1144/gsl.sp.2001.190.01.14. ISBN 978-1-86239-093-5. LCCN 2003464816. OCLC 48570033. S2CID 130092094.
- Дарвин, Чарльз (1887). Дарвин, Фрэнсис (ред.). The Life and Letters of Charles Darwin, Including an Autobiographical Chapter. 3 (3-е изд.). Лондон: Джон Мюррей. OCLC 834491774.
- Davies, Geoffrey F. (2007). «Chapter 2.3 Dynamics of the Hadean and Archaean Mantle». In van Kranendonk, Martin J.; Smithies, R. Hugh; Bennett, Vickie C. (eds.). Earth’s Oldest Rocks. События в геологии докембрия. 15. Амстердам; Бостон: Эльзевир. С. 61–73. Дои:10.1016/S0166-2635(07)15023-4. ISBN 978-0-444-52810-0. LCCN 2009525003.
- Дэвис, Пол (1999). The Fifth Miracle: The Search for the Origin of Life. Лондон: Книги о пингвинах. ISBN 978-0-14-028226-9.
- Докинз, Ричард (1996). Слепой часовщик (Reissue with a new introduction ed.). Нью-Йорк: W.W. Нортон и компания. ISBN 978-0-393-31570-7. LCCN 96229669. OCLC 35648431.
- Dawkins, Richard (2004). Сказка предков: паломничество к заре эволюции. Бостон: Houghton Mifflin. ISBN 978-0-618-00583-3. LCCN 2004059864. OCLC 56617123.
- Добелл, Клиффорд (1960) [Originally published 1932; Нью-Йорк: Harcourt, Brace & Company ]. Antony van Leeuwenhoek and His ‘Little Animals’. Нью-Йорк: Dover Publications. LCCN 60002548.
- Дайсон, Фриман (1999). Истоки Жизни (Пересмотренная ред.). Cambridge, UK; Нью-Йорк: Издательство Кембриджского университета. ISBN 978-0-521-62668-2. LCCN 99021079.
- Eigen, M.; Schuster, P. (1979). The Hypercycle: A Principle of Natural Self-Organization. Берлин; Нью-Йорк: Springer-Verlag. ISBN 978-0-387-09293-5. LCCN 79001315. OCLC 4665354.
- Fesenkov, V.G. (1959). «Some Considerations about the Primaeval State of the Earth». В Oparin, A.I.; и другие. (ред.). The Origin of Life on the Earth. I.U.B. Symposium Series. 1. Edited for the International Union of Biochemistry by Frank Clark and R.L.M. Synge (English-French-German ed.). Лондон; Нью-Йорк: Pergamon Press. ISBN 978-1-4832-2240-0. LCCN 59012060. Получено 3 июн 2015. International Symposium on the Origin of Life on the Earth (held at Moscow, 19–24 August 1957)
- Хейзен, Роберт М. (2005). Genesis: The Scientific Quest for Life’s Origin. Вашингтон, округ Колумбия: Джозеф Генри Пресс. ISBN 978-0-309-09432-0. LCCN 2005012839. OCLC 60321860.
- Хаксли, Томас Генри (1968) [Originally published 1897]. «VIII Biogenesis and Abiogenesis [1870]». Discourses, Biological and Geological. Collected Essays. VIII (Перепечатка ред.). Нью-Йорк: Гринвуд Пресс. LCCN 70029958.
- Kauffman, Stuart (1993). The Origins of Order: Self-Organization and Selection in Evolution. Нью-Йорк: Oxford University Press. ISBN 978-0-19-507951-7. LCCN 91011148. OCLC 23253930.
- Kauffman, Stuart (1995). At Home in the Universe: The Search for Laws of Self-Organization and Complexity. Нью-Йорк: Издательство Оксфордского университета. ISBN 978-0-19-509599-9. LCCN 94025268.
- Klyce, Brig (22 January 2001). Kingsley, Stuart A.; Бхатал, Рагбир (ред.). Panspermia Asks New Questions. The Search for Extraterrestrial Intelligence (SETI) in the Optical Spectrum III. 4273. Bellingham, WA: SPIE. Дои:10.1117/12.435366. ISBN 0-8194-3951-7. LCCN 2001279159. Получено 9 июн 2015. Слушания SPIE, проходившие в Сан-Хосе, Калифорния, 22–24 января 2001 г.
- Лейн, Ник (2009). Восхождение жизни: 10 великих изобретений эволюции (1-е американское изд.). Нью-Йорк: W.W. Нортон и компания. ISBN 978-0-393-06596-1. LCCN 2009005046. OCLC 286488326.
- Ланкенау, Дирк-Хеннер (2011). «Два мира РНК: к происхождению репликации, генов, рекомбинации и ремонта». В Эгеле, Ричард; Ланкенау, Дирк-Хеннер; Мулкиджанян, Армен Ю. (ред.). Истоки жизни: изначальная самоорганизация. Гейдельберг: Springer. Дои:10.1007/978-3-642-21625-1. ISBN 978-3-642-21624-4. LCCN 2011935879. OCLC 733245537.
- Леннокс, Джеймс Г. (2001). Философия биологии Аристотеля: исследования истоков наук о жизни. Кембриджские исследования в области философии и биологии. Кембридж, Великобритания; Нью-Йорк: Издательство Кембриджского университета. ISBN 978-0-521-65976-5. LCCN 00026070.
- Мичод, Ричард Э. (1999). Дарвиновская динамика: эволюционные переходы в приспособленности и индивидуальности. Принстон, штат Нью-Джерси: Princeton University Press. ISBN 978-0-691-02699-2. LCCN 98004166. OCLC 38948118.
- Опарин, А. (1953) [Первоначально опубликовано в 1938 году; Нью-Йорк: Компания Macmillan ]. Происхождение жизни. Перевод и новое введение Сергия Моргулиса (2-е изд.). Минеола, Нью-Йорк: Dover Publications. ISBN 978-0-486-49522-4. LCCN 53010161.
- Оргел, Лесли Э. (1987). «Эволюция генетического аппарата: обзор». Эволюция каталитической функции. Симпозиумы Колд-Спринг-Харбор по количественной биологии. 52. Колд-Спринг-Харбор, Нью-Йорк: Лаборатория Колд-Спринг-Харбор. С. 9–16. Дои:10.1101 / SQB.1987.052.01.004. ISBN 978-0-87969-054-0. OCLC 19850881. PMID 2456886. «Труды симпозиума, проведенного в лаборатории Колд-Спринг-Харбор в 1987 г.»
- Рэйвен, Питер Х.; Джонсон, Джордж Б. (2002). Биология (6-е изд.). Бостон: Макгроу-Хилл. ISBN 978-0-07-112261-0. LCCN 2001030052. OCLC 45806501.
- Росс, Александр (1652). Arcana Microcosmi. Книга II. Лондон. Получено 7 июля 2015.
- Шапиро, Роберт (1987). Происхождение: руководство скептика по созданию жизни на Земле. Торонто; Нью-Йорк: Bantam Книги. ISBN 978-0-553-34355-7.
- Шелдон, Роберт Б. (22 сентября 2005 г.). Гувер, Ричард Б.; Левин, Гилберт В.; Розанов, Алексей Юрьевич .; Гладстон, Г. Рэндалл (ред.). Историческое развитие различия между био- и абиогенезом (PDF). Астробиология и планетарные миссии. 5906. Беллингхэм, Вашингтон: SPIE. Дои:10.1117/12.663480. ISBN 978-0-8194-5911-4. LCCN 2005284378. Получено 13 апреля 2015. Слушания SPIE, проходившие в Сан-Диего, Калифорния, 31 июля — 2 августа 2005 г.
- Тиндалл, Джон (1905) [Первоначально опубликовано в 1871 году; Лондон; Нью-Йорк: Longmans, Green & Co .; Д. Эпплтон и компания ]. Фрагменты науки. 2 (6-е изд.). Нью-Йорк: ПФ. Кольер и сыновья. OCLC 726998155. Получено 6 июн 2015.
- Вартанян, Арам (1973). «Спонтанное зарождение». В Винер, Филип П. (ред.). Словарь истории идей. IV. Нью-Йорк: Сыновья Чарльза Скрибнера. ISBN 978-0-684-13293-8. LCCN 72007943.
- Воет, Дональд; Воет, Джудит Г. (2004). Биохимия. 1 (3-е изд.). Нью-Йорк: Джон Вили и сыновья. ISBN 978-0-471-19350-0. LCCN 2003269978.
- Вудворд, Роберт Дж., Изд. (1969). Наш удивительный мир природы: его чудеса и тайны. Плезантвилл, Нью-Йорк: Reader’s Digest Association. ISBN 978-0-340-13000-1. LCCN 69010418.
- Ярус, Михаил (2010). Жизнь из мира РНК: Предок внутри. Кембридж, Массачусетс: Издательство Гарвардского университета. ISBN 978-0-674-05075-4. LCCN 2009044011.
- Arrhenius, Gustaf O .; Продажи, Брайан К .; Mojzsis, Стивен Дж .; и другие. (21 августа 1997 г.). «Энтропия и заряд в молекулярной эволюции — случай фосфата» (PDF). Журнал теоретической биологии. 187 (4): 503–522. Дои:10.1006 / jtbi.1996.0385. PMID 9299295. Архивировано из оригинал (PDF) 19 мая 2017 г.. Получено 30 июн 2015.
- Кавальер-Смит, Томас (Июнь 2006 г.). «Клеточная эволюция и история Земли: застой и революция». Философские труды Королевского общества B. 361 (1470): 969–1006. Дои:10.1098 / rstb.2006.1842. ЧВК 1578732. PMID 16754610.
- Fernando, Chrisantha T .; Роу, Джонатан (7 июля 2007 г.). «Естественный отбор в химической эволюции». Журнал теоретической биологии. 247 (1): 152–167. CiteSeerX 10.1.1.129.8556. Дои:10.1016 / j.jtbi.2007.01.028. PMID 17399743.
- Гросс, Майкл (19 декабря 2016 г.). «Как жизнь может возникнуть из химии». Текущая биология. 26 (24): R1247 – R1249. Дои:10.1016 / j.cub.2016.12.001.
- Хорган, Джон (Февраль 1991 г.). «В начале.». Scientific American. 264 (2): 116–125. Bibcode:1991SciAm.264b.116H. Дои:10.1038 / scientificamerican0291-116.
- Игнатов, Игнат; Мосин, Олег В. (2013). «Моделирование возможных процессов происхождения жизни и живой материи в горячих минералах и морской воде с дейтерием». Журнал наук об окружающей среде и Земле. 3 (14): 103–118. Получено 29 июн 2015.
- Джортнер, Джошуа (Октябрь 2006 г.). «Условия возникновения жизни на ранней Земле: краткое содержание и размышления». Философские труды Королевского общества B. 361 (1474): 1877–1891. Дои:10.1098 / rstb.2006.1909. ЧВК 1664691. PMID 17008225.
- Клотц, Ирэн (24 февраля 2012 г.). «Жизнь началась в пруду, а не в океанах?». Новости открытия. Сильвер Спринг, Мэриленд: Discovery Communications. Получено 29 июн 2015.
- Институт астробиологии НАСА: Харрисон, Т. Марк; МакКиган, Кевин Д.; Мойзсис, Стивен Дж. «Ранняя среда и жизнь Земли: когда Земля стала пригодной для проживания?». Архивировано из оригинал 17 февраля 2012 г.. Получено 30 июн 2015.
- Специализированный центр исследований и подготовки специалистов по экзобиологии НАСА: Аррениус, Густав О. (11 сентября 2002 г.). «Аррениус». Архивировано из оригинал 21 декабря 2007 г.. Получено 30 июн 2015.
- «Физико-химические основы жизни». Что такое жизнь. Спринг-Вэлли, Калифорния: Лукас К. Бюлер. Получено 27 октября 2005.
- Питч, Стефан; Кришнамурти, Раманараянан; Аррениус, Густав О. (6 сентября 2000 г.). «Концентрация простых альдегидов сульфитсодержащими двухслойными гидроксидными минералами: значение для биопоэза». Helvetica Chimica Acta. 83 (9): 2398–2411. Дои:10.1002 / 1522-2675 (20000906) 83: 9 <2398 :: AID-HLCA2398> 3.0.CO; 2-5. PMID 11543578.
- Понс, Мари-Лор; Quitté, Ghylaine; Фудзи, Тошиюки; и другие. (25 октября 2011 г.). «Ранние архейские серпентиновые грязевые вулканы в Исуа, Гренландия, как ниша для ранней жизни». Proc. Natl. Акад. Sci. СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ. 108 (43): 17639–17643. Bibcode:2011PNAS..10817639P. Дои:10.1073 / pnas.1108061108. ЧВК 3203773. PMID 22006301.
- Рассел, Майкл Дж .; Холл, A.J .; Кэрнс-Смит, Александр Грэм; и другие. (10 ноября 1988 г.). «Подводные горячие источники и зарождение жизни». Природа. 336 (6195): 1569–1572. Bibcode:1988Натура.336..117R. Дои:10.1038 / 336117a0. S2CID 45788156.
- Шок, Эверетт Л. (25 октября 1997 г.). «Высокотемпературная жизнь без фотосинтеза как модель Марса» (PDF). Журнал геофизических исследований. 102 (E10): 23687–23694. Bibcode:1997JGR … 10223687S. Дои:10.1029 / 97je01087. PMID 11541237.
дальнейшее чтение
- Тим Флэннери, «In the Soup» (рецензия Майкла Маршалла, The Genesis Quest: гении и чудаки в пути, чтобы раскрыть истоки жизни на Земле, University of Chicago Press, 360 стр.), Нью-Йоркское обозрение книг, т. LXVII, нет. 19 (3 декабря 2020 г.), стр. 37–38.
внешняя ссылка
Послушайте эту статью
(4 части, 129,1 мегабайт)
![]()
Эти аудиофайлы были созданы на основе редакции этой статьи от 13 июня 2012 г., и не отражают последующие правки.
(Аудио помощь · Больше устных статей)
- «Изучение истоков жизни: виртуальная выставка». Изучение истоков жизни: виртуальная выставка. Округ Арлингтон, Вирджиния: Национальный фонд науки. Получено 2 июля 2015.
- «Геохимическое происхождение жизни Майкла Дж. Рассела и Аллана Дж. Холла». Глазго, Шотландия: Университет Глазго. 13 декабря 2008 г.. Получено 2 июля 2015.
- Мэлори, Марсия. «Как зародилась жизнь на Земле». Факты о Земле. Получено 2 июля 2015.
- Истоки жизни, Дискуссия BBC Radio 4 с Ричардом Докинзом, Ричардом Корфилдом и Линдой Партридж (В наше время, 23 сентября 2004 г.)
- Очерк Уинстона Черчилля об эволюции (1939-1950-е годы)

Привет! Нашёл диагностический адаптер Thinkdiag TKD01 и с помощью Diagzone выполнил сканирование. Вылезла следующая ошибка:
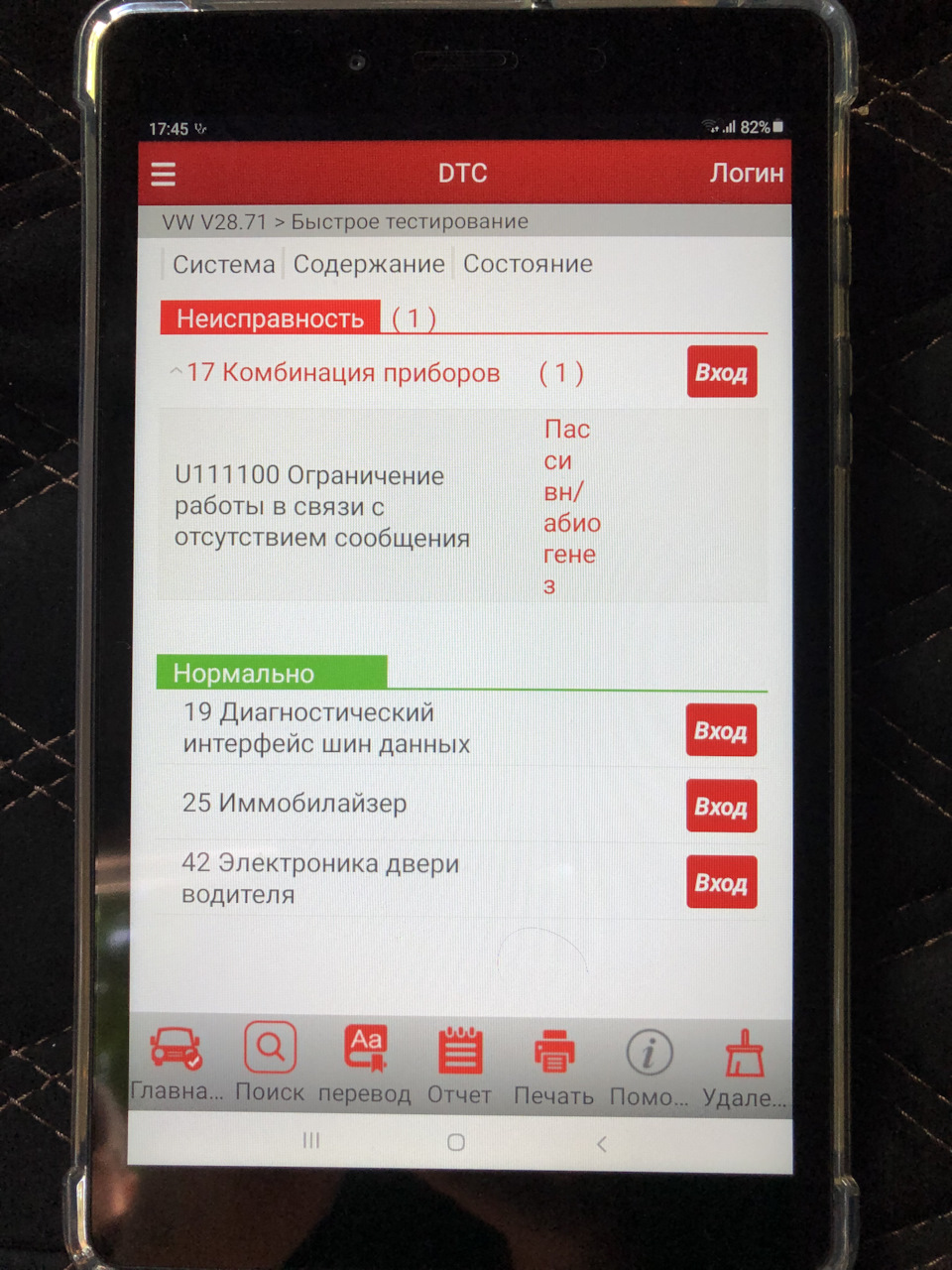
Полный размер
Удалилась без проблем и пока не появлялась. Кто подскажет, о чем вообще речь? Перевод программы оставляет желать лучшего. На приборке ошибок не было замечено. После быстрого поиска по форумам было найдено, что это может быть связано с плохим контактом проводов, окислением разъёмов и пр.

Полный размер
Запчасти на фото: 5K0920871A
Пробег: 262 000 км
Войдите или зарегистрируйтесь, чтобы писать комментарии, задавать вопросы и участвовать в обсуждении.
Обновлено: 05.06.2023
При диагностике выбило такую ошибку, диагносты пожимают плечами, разшифруйте пожалуйста ошибку( была проблема с задним датчиком дорожного просвета, сняли его и почистили, после пытались адаптировать и выдало такую ошибку, с датчиком проблема осталась, датчик воще подлежит ремонту или менять на новый?)
Я здесь живу
Vaxa20
Самоделкин
Датчик только под замену он электроникой напичкан, ошибка — не запрограмированно рабочее положение, слетела калибровка подвески ввиду ее адаптации с неиправным датчиком судя по всему.
mishanka80
Мастер советчик
mishanka80
Мастер советчик
Датчик только под замену он электроникой напичкан, ошибка — не запрограмированно рабочее положение, слетела калибровка подвески ввиду ее адаптации с неиправным датчиком судя по всему.
Я здесь живу
Почитай вот здесь,думаю все найдешь,все разжеванно.
mishanka80
Мастер советчик
Спасибо, все так и делали, только в канале 2-3-4— не открывается.Логин принимает и открывает только 1 и 10 канал.Ошибку удалить не получается, советовали чтобы авто типа самообучился, тоесть поездить в разных режимах назад, вперед,влево и вправо, всеравно ошибка остается.Что делать?
mishanka80
Мастер советчик
Да и поповоду датчика- он же не механический а индуктивный и вроде изнашиватся там нечему?Он работает но в определенных положениях сигнал проподает, проводкуи разьем смотрел.
Я здесь живу
Странно что тебя к трем колесам она не допускает,а кодировка блока случаем не слетела,глянь че там в кодировке стоит.
Vaxa20
Самоделкин
Датчик там не индуктивный, лично разберал, с алроуда, там с датчиком холла + микруха на выходе стоит хотя по схеме вообще переменик нарисован! По калибровке, сочуствую, операция не сложная но нужны калибры в заднюю подвеску, раз калибровал без калибров самое настоящие мучение (в виду того что ЭБУ подвески заменили) на домкратах выводил. в краце чтобо схватило, надо выставить ровно машину опустить машину на калибры в моем случае на домкраты(позиция для дороги), разница в показания датчиков не белее 5 ед ( инача не возьмет) в противном случаем лезем выводим значения датчиком там помоему есть не большая регулировка, при этих действиях зад калибруется на ура, а вот перед это я уж не знаю какая там у тебя машина мне попалась что при полностью спущеном состоянии подушек она кривая была и замучился ее выставлять положения чтоб она схватила эти значения. Я делал все по эльзе там все хорошо написано но ввиду кривых машин и отсутствия спец инструмента убил пол дня на нее. а так по хорошему на калибровку если все испарвно 10 мин достаточно!
И еще проверь действительно кодировку блока как говорит аКа4, не снесли ее тебе попутно случаем
ten70
Оракул
Для начала можно сбросить настройки по 00 каналу.
mishanka80
Мастер советчик
ten70
Оракул
Была подобная проблема ничего не помогало, тряхнули подвеску на подъемнике- заработало.
Списали на залипший датчик.
mishanka80
Мастер советчик
mishanka80
Мастер советчик
Для начала можно сбросить настройки по 00 каналу.
Я здесь живу
Техническое описание и расшифровка ошибки P2261
Сохраненный код неисправности P2261 указывает, что модуль управления (PCM) обнаружил проблему в системе турбокомпрессора или нагнетателя. В данном случае, обнаружена механическая неисправность перепускного клапана.

В системах с принудительной индукцией, давление на входе должно изменяться и регулироваться в соответствии с необходимой мощностью на данный момент. Для этого используется регулирующий клапан наддува. Который контролируется и управляется модулем управления двигателем, чтобы обеспечить двигатель идеальной топливно-воздушной смесью.
Это происходит за счет механической регулировки лопаток турбонагнетателя/нагнетателя. Лопатки отвечают за регулировку количества наддува, который подает воздух в камеру сгорания. Если PCM теряет контроль над наддувом, это приводит к неустойчивой работе двигателя.
Код неисправности P2261 активируется, когда модуль управления двигателем обнаруживает, что произошла неисправность перепускного клапана турбокомпрессора. В данном случае указывается конкретная проблема, а именно механическая.
После чего, контроллер включает контрольную лампу двигателя (MIL). А ошибка записывается в память, как неисправность в системе управления наддувом.
Для включения контрольной лампы MIL на панели приборов, может потребоваться несколько циклов с отказом.
Симптомы неисправности
Также они могут проявляться как:
Уровень серьезности от среднего до высокого, в зависимости от симптомов. Ошибку P2261 игнорировать не стоит, так как вы рискуете повредить внутренние компоненты двигателя. Поэтому в ваших интересах, как можно скорее устранить неисправности в системе принудительной индукции.
Причины возникновения ошибки
Код P2261 может означать, что произошла одна или несколько следующих проблем:
- Механическая неисправность перепускного клапана.
- Коррозия, вызывающая высокое сопротивление в разъемах или контактах.
- Изношенная или замкнутая проводка.
- Забитый или поврежденный воздушный фильтр.
- Утечка воздуха в вакуумной системе (часто утечка обнаруживается в вакуумном баке).
- Иногда причиной является неисправный модуль PCM.
Как устранить или сбросить код неисправности P2261
Некоторые предлагаемые шаги для устранения неполадок и исправления кода ошибки P2261:
- Считайте все сохраненные данные и коды ошибок с помощью сканера OBD-II.
- Очистите коды ошибок с памяти компьютера и проведите тест-драйв автомобиля, чтобы выяснить, появляется ли ошибка P2261 снова.
- Проверьте перепускной клапан.
- Проверьте вакуумные линии на предмет утечек.
- Осмотрите воздушный фильтр, возможно, он требует замены.
- Протестируйте разъем и проводку на наличие повреждений, ослабления и коррозии.
- Проверьте модуль PCM.
Диагностика проблем и решение проблем
Турбины обычно вращаются с невероятной скоростью от 100 000 до 150 000 оборотов в минуту. Они не терпимы к дисбалансу или отсутствию чистого масла в подшипнике.
Процесс диагностики при ошибке P2261 лучше начать с наиболее распространенных проблем с турбонаддувом. Для их проведения понадобятся инструменты, такие как вакуумметр и циферблатный индикатор.
Проверка вакуума
Убедитесь, что двигатель работает должным образом, без пропусков зажигания и кодов, относящихся к неисправному датчику детонации. Затем, проверьте герметичность хомутов на выходе турбонагнетателя, промежуточного охладителя, а также корпусе дроссельной заслонки.
Осмотрите впускной коллектор на предмет утечек любого рода, включая вакуумные шланги. Снимите рычаг с перепускной заслонки. Вручную управляйте клапаном, ища заедание клапана, вызывающее падение наддува.
При поиске утечки, обратите особое внимание на вакуумный бак, именно он является частой причиной данной проблемы.
Замер давления наддува, осмотр турбонагнетателя
Найдите вакуум без отверстий во впускном коллекторе и установите вакуумметр. Запустите двигатель. На холостом ходу двигатель должен иметь разрежение 1-1.5 атмосферы. Если меньше 1 атмосферы, значит каталитический нейтрализатор неисправен и не позволяет наращивать наддув.
Быстро разгоните двигатель до 5000 об / мин и отпустите дроссельную заслонку, наблюдая за вакуумметром, показывающим давление наддува. Если давление наддува поднимается выше 1.3 атмосферы, значит плохой перепускной клапан.
Заглушите двигатель и дайте ему остыть. Снимите выпускной шланг турбины и загляните внутрь, чтобы убедиться, что лопасти не задевают корпус. Ищите погнутые или отсутствующие лопасти или масло в турбонагнетателе. Вращайте лопасти вручную, ищите сопротивление, указывающее на неисправность турбонагнетателя.
Осмотрите маслопроводы от блока цилиндров к центральному подшипнику и возвратный трубопровод от подшипника к масляному поддону на предмет утечек. Установите циферблатный индикатор на выходной патрубок турбины, прокрутите вал турбины. Если осевой люфт превышает 0,003, центральный подшипник неисправен.
Проводка и датчик давления
Если после всех проверок проблемы не обнаружены, но код P2261 не пропал, необходимо проверить электрические компоненты. Нужно протестировать датчик и связанные с ними цепи.
Отсоедините жгут проводов и протестируйте цепь управления. Убедитесь, что в цепи питания присутствует 12 вольт. Они должны показываться при включенном зажигании. Если их нет, или если они присутствуют при выключенном зажигании, отремонтируйте проводку от PCM к датчику.
Проверьте надежность заземления. Подключите контрольную лампу к плюсу аккумуляторной батареи, а другим концом коснитесь цепи заземления датчика. Если контрольная лампа не загорается, это указывает на неисправную цепь.
На каких автомобилях чаще встречается данная проблема
Проблема с кодом P2261 может встречаться на различных машинах, но всегда есть статистика, на каких марках эта ошибка присутствует чаще. Вот список некоторых из них:
- Audi (Ауди а4)
- Buick (Бьюик Энкор)
- Chevrolet (Шевроле Круз)
- Citroen
- Honda
- Hyundai (Хендай Велостер, Генезис)
- Kia
- Mercedes
- Opel (Опель Астра, Инсигния)
- Peugeot (Пежо 308, 3008)
- Volkswagen (Фольксваген Тигуан)
С кодом неисправности Р2261 иногда можно встретить и другие ошибки. Наиболее часто встречаются следующие: P0234, P0299, P0300, P0321, P2187, P2262, P2263, P226B, P3081.
Конструктивно самолет не сложнее современной легковушки. Но в кабине пилота, бесчисленное количество кнопок и тумблеров, а на торпеде автомобиля их нет. Почему так?
Поэтому автопроизводители предусматривают систему контрольных ламп на приборном щитке, которые сигнализируют о проблемах в технике или электронике. Одним из таких индикаторов служит значок EPC. Что он значит? Чего бояться и куда бежать? Об этом ниже.
Что означает значок ЕРС на приборной панели Ауди
Обратите внимание! У многих немецких автомобилях при повороте ключа зажигания индикатор EPC загорается на пару секунд, после чего гаснет. Это значит, что система работает нормально, а вот если горит постоянно — это повод обратиться в сервис.

Ошибка EPC на Audi A3
В каких случаях он загорается
Индикатор EPC загорается при обнаружении любой проблемы, связанной с бортовой электроникой и реакцией датчиков на неисправности механического характера. Зачастую, контрольная лампа свидетельствует о:
Определить проблему на глаз не получится. При подключении компьютера или смартфона система сама покажет номер ошибки. После чего остается посмотреть, что она значит.
Бывают и не стандартные ситуации. Например, EPC ошибка на Ауди показывает неисправность датчика давления в картере. Но на деле с ним все в порядке. Оказалось, что была плохо закручена крышка горловины после долива масла (нужно до щелчка). В системе шел подсос воздуха и давление в картере оказалось ниже нормы, что привело к срабатыванию контрольной лампы. Но это исключительный случай.
Безопасно ли ездить с таким индикатором

Учитывая, что контрольная лампа может гореть даже от банального перегорания лампочки или реле, то в каких-то случаях можно ездить и не беспокоиться. Например, неисправный датчик дождя для многих не станет весомым поводом обращаться в сервис и платить от 1000 руб. за диагностику.
Ошибки EPC по моделям
Ошибок и неисправностей, о которых сигнализирует EPC — сотни. Всех, естественно, не перечислить. Все рассчитано на то, чтобы автовладелец обратился в сервисный центр. Но, как говорится, у каждой модели есть свои характерные проблемы, на которые жалуются владельцы на профильных форумах. Ниже краткая выжимка по некоторым моделям бренда.
Индикатор EPC на приборной панели у владельцев Ауди Q5 часто выдает следующие ошибки:
Что интересно, у многих владельцев Audi Q5 EPS загорался, когда Ингольштадт выкатывал новую прошивку, которую предлагалось установить в добровольно-принудительном порядке.

Ошибка EPC на приборке Audi Q5
- Отсутствие сигнала от турбины гидротрансформатора АКПП. Проблема решается заменой масла, датчика температуры или корпуса клапана. По ситуации;
- Ошибочный сигнал от датчика оборотов двигателя. Потребуется его замена или визуальный осмотр на наличие коррозийных проводов;
- Проблема с передаточным отношением в КПП. Ремонт зависит от характера поломки. Начиная от замены проводов и масляного насоса, заканчивая полной заменой коробки.
Также, конкретно на Ауди А4 B8 ошибка P039в в EPC свидетельствует о высоком давлении в первом цилиндре. Возможна проблема с турбокомпрессором. Распространенная болячка.
Как убрать ошибку УЗС на Ауди А4 1.8Т, смотрите в видеоролике ниже:
По Ауди А6 автомобилисты также часто спрашивают по таким ошибкам, как:
- Неисправность в датчике скорости. Ремонт может не ограничится парой проводов или самим датчиком. Периодически приходится менять весь модуль ECM;
- Высокая температура модуля управления мотором. При ремонте проверяется заземление и предохранители. При необходимости модуль меняется;
- Ненормированное напряжение во входной цепи уровня топлива. Ремонт может ограничится заменой проводов и соединителей, либо заменой модуля РСМ.
Также, если брать конкретно Ауди A6 C5, в EPC частая ошибка P1676, которая свидетельствует об обрыве цепи контрольной лампы левого стоп-сигнала. Ремонтируется путем поиска обрыва, проверки изоляции и замены коррозийных проводов.
Резюмируя, ошибок в системе EPC сотни и каждая сигнализирует об определенной проблеме. Это значительно упрощает ремонт высокотехнологичных авто и помогает вовремя предупредить серьезные неисправности двигателя и КПП, пока стоимость ремонта не превысила стоимость самой машины.
Ошибка EPC и чек двигателя на автомобилях VAG — тема данного видео:
В регистраторе событий блока управления двигателя содержатся следующие события:
«P1127: ряд1 — адаптация смеси (под нагрузкой), переобогащение смеси»
«P1129: ряд2 — адаптация смеси (под нагрузкой), переобогащение смеси»
В некоторых случаях неблагоприятный режим движения может привести к повышенному содержанию топлива в моторном масле и/или к затруднённому удалению топлива из моторного масла.
В случае рекламации выполнить следующие работы:
Выбрать в блоке управления двигателя следующие измеряемые величины:
Долговременная адаптация смеси, ряд 1
Долговременная адаптация смеси, ряд 2
Если значение для одного ряда или для обоих рядов находится в диапазоне ~ -10…-20%, в моторном масле предположительно присутствует топливо.
Проверить герметичность топливных форсунок. Для этого двигатель должен быть прогрет до рабочей температуры. Выбрать в блоке управления двигателя следующую измеряемую величину:
Давление топлива в контуре высокого давления, фактическое
Заглушить двигатель и проверить изменение давления по указанной измеряемой величине. Если форсунки герметичны, значение измеряемой величины возрастёт в течение нескольких минут (повышение давления из-за аккумулирования тепла в топливной рампе).
Если давление топлива повышается, результат проверки герметичности положительный, негерметичность форсунок исключается.
Если приближается срок очередного технического обслуживания, идеальным решением была бы замена моторного масла. В противном случае необходимо выполнить пробную поездку при температуре моторного масла более 80°C в течение не менее 20 минут. Затем повторно выбрать измеряемые величины Долговременная адаптация смеси, ряд 1/ряд 2. Эти значения после пробной поездки должны быть около 0%.
Решение технических проблем
Если вы не нашли информацию по своему автомобилю — посмотрите ее на автомобили построенные на платформе вашего авто.
С большой долей вероятности информация по ремонту и обслуживанию подойдет и для Вашего авто.
Если открывает только 1 и 10 канал, значит логин не проходит.
ВАГКОМОМ нужно пробовать подключаться, в добавок вот что написано:
; VAG-COM Measuring Block Labels file.
; Module 34 — Level Control
; This module seems very timing sensitive, be sure to close any unneeded background tasks
mishanka80
Мастер советчик
Да нет, если ввожу неправильный логин нет ни одного откр. канала, вожу правильный появляются 0,1, и 10.Что скажете по ошибке, незря же она выскочила?
ten70
Оракул
01437 — Control Position not Learned
Possible Symptoms
* Malfunction Indicator Light (MIL) ON
* Control Position not learned
* Check Control Module/System Adaptation and/or Basic Setting
Для начала нужно выполнить некоторые условия:
* Transmission in park/neutral.
* Start the vehicle, keep the engine running and do not switch off the ignition.
* Doors have to stay closed, otherwise the system is not ready.
Попробуй ввести логин в :
mishanka80
Мастер советчик
01437 — Control Position not Learned
Possible Symptoms
* Malfunction Indicator Light (MIL) ON
* Control Position not learned
* Check Control Module/System Adaptation and/or Basic Setting
Для начала нужно выполнить некоторые условия:
Попробуй ввести логин в :
ten70
Оракул
Чтобы войти в базовые установки нужно сперва
КПП перевести на нейтралку/паркинг.
Завести двигатель и не выключать.
Закрыть все двери в авто.
Иначе в базовые установки не войдешь.
mishanka80
Мастер советчик
Чтобы войти в базовые установки нужно сперва
КПП перевести на нейтралку/паркинг.
Завести двигатель и не выключать.
Закрыть все двери в авто.
Иначе в базовые установки не войдешь.
ten70
Оракул
Ошибка называется — система управления высотой не обучена.
Я вообще не понимаю нахрена датчик трогали.
Попробуй ввести логин в 16 кнопочке если она активна.
ten70
Оракул
Попробуй начать адаптацию с 1 канала, потом 2 и тд и тп.
При адаптации ждать нужно, пока она закончится.
mishanka80
Мастер советчик
Я здесь живу
mishanka80
Мастер советчик
Ето понятно с датчиком, я же говорю что подкидывал датчик другой и ошибка не пропадала.Воще все началось с того что задняя левая сторона чуть опущена была(там где стоит проблемный датчик).Сняли его и почистили, он заработал без проблем(в ручную тягу перемещал и показания не прыгали).Потом начали адаптацию— было две ошибки связаные с фарами и датчиком, их удалил.Вел логин и вошол в адаптацию, открыл 1затем 2 и тд. в 5 поставил 1. Когда вышел и снова вошол в пневматику чтобы проверить наличие ошибок выскочила ета ошыбка.Далее обратился за помощю сюда.Подвеска работает нормально(поднимает и опускает) но на панели постоянно горит лампа неисправности пневмат.
ten70
Оракул
Какие значения выставлял?
The default values:
Audi A6 (4F) (PR-1BK): 386 mm (front) and 384 mm (rear)
Audi A6 (4F) Allroad (PR-1BY): 388 mm (front) and 380 mm (rear)
Audi A8 (4E) standard suspension (PR-1BK): 416 mm (front) and 398 mm (rear)
Audi A8/S8 (4E) sport suspension (PR-2MA/2MB): 396 mm (front) and 378 mm (rear)
Читайте также:
- B1618 ошибка тойота прадо
- Старлайн автозапуск как настроить автозапуск по температуре
- Рено аркана автозапуск с телефона своими руками схема
- C003a ошибка abs ford
- Ошибки лямбда зонда бмв е39
Опубликовано: 05.06.2023
Основные проблемы моторов TFSI:
_______________________________________________________
1. Неисправность перепускного клапана (байпас, вестгейт)
2. PCV (неисправность клапана вентиляции картерных газов)
3. Износ толкателя ТНВД
4. Неисправность ТНВД
5. Не работает моторчик впускных заслонок
6. Пропуски зажигания
7. Неисправность датчика давления топлива
8. Цепной звук на холодную.
9. Неисправность клапана рециркуляции N80 (EVAP)
10. Залегание колец из-за странной их конструкции.
11. OIL PRESSURE
_______________________________________________________
1. Перепускной клапан на турбине (байпас, вестгейт).
Симптомы:
— Неисправный клапан травит давление.
— Давление можно посомтреть ВАГ-КОМом, группа 115. Будет видно резкое снижение давления наддува.
-Давление при этом примерно 0,5-0,6бар. Может варьироваться.
-Возможно, будет ошибки вида: 000665 — Boost Pressure Regulation: Control Range Not Reached
p0299 — 002 — Lower Limit Exceeded — Intermittent
—Ошибка P1297
— Как вариант, можно снять и осмотреть перепускной клапан на предмет целостности диафрагмы. Крепится он на трех болтах, к холодной части турбины. Мембрана будет частично или полностью повреждена.
Решение: На сегодняшний день есть много вариантов. Самые распространенные: замена на улучшенный стандартный (06H145710D).____________________________________________________________________________________
2. Клапан вентиляции картерных газов.
Изначально система расчитана на всасывания газов из картера во впускной коллектор (ресивер), когда мотор не под давлением (холостой ход, вакуум). Если клапан не исправен, при повышении давления газы из коллектора обратно не уходят в картер.
Симптомы:
— Небольшое падение давления (до 0.1 бара в среднем)
-Возможно выдавливание масла через крышку маслозаливной горловины или свечные колодцы.
— Возможно небольшое увеличение расхода топлива.
— Засорение заднего патрубка (клапана) вентиляции приводит к выдавливанию масла в районе перепускного клапана, возможен немного синий дым при полном газе.
Решение: Замена клапана.
Детали, для замены клапана:
06F 129 101 N (сам клапан, есть разные ревизии)
06F 103 483 E (прокладка).____________________________________________________________________________________
3. Износ толкателя ТНВД.
Износ происходит из-за соприкосновения кулачка с толкателем ТНВД. Повышенный износ возможен из-за повреждения толкателя ТНВД.
Симптомы:
— Пропуски (провалы) по топливу, особенно при полном газе
— Пропуски по топливу могут быть как короткими, так и длинными на высоких оборотах. В особо «тяжелых» случаях возможно загорание ЧекЭнджина и перехода мозга в аварийный режим ( максимум 4500обмин), с падением давления наддува.
— Ошибка P0087 Fuel Rail / System Pressure — Too Low
— Ошибка P1093 Fuel Trim 2, Bank 1 Malfunction
— Ошибка P2293 Fuel Pressure Regulator 2 Performance
Решение: Замена толкателя ТНВД (если необходимо), шлифовка кулачка распредвала, либо замена распредвала.____________________________________________________________________________________
4. Неисправность ТНВД.
Симптомы:
— Возможные провалы при полном газе.
— Возможны ошибки : 004767 — Low-Pressure Fuel Pressure Regulation: Pressure too High
P129F — 001 — Upper Limit Exceeded — MIL ON
Решение: Проверить кулачок ТНВД, при необходимости заменить.____________________________________________________________________________________
5. Не работает моторчик впускных заслонок.
Неработающие заслонки (читай моторчик заслонок) одна из самых распространенных проблем на TFSI.
Основной принцип работы показан на картинке:
Симптомы:
Возможно появление ошибок:
-012599 — Intake Manifold Runner Control: Basic Setting not Completed
P3137 — 001 — Upper Limit Exceeded – Intermittent
-012691 — Intake Manifold Runner Control: Open Stop outside of Valid Range
P3193 — 002 — Lower Limit Exceeded — Intermittent
-008196 — Intake Manifold Flap; Bank 1: Stuck Open
P2004 — 008 — Implausible Signal — Intermittent
— 012600 — Intake Manifold Runner Control: Regulation Deviation
P3138 — 008 — Implausible Signal — MIL ON
ОШИБКИ:
P1098 — Датчик положения заслонок впускных каналов(потенциометр) G336: обрыв цепи/короткое замыкание на плюс.
P3193 — Заслонки впускных каналов: Упор в положении ОТКР. за пределами действительного диапазона.
Решение: Замена моторчика впускных заслонок.____________________________________________________________________________________
6. Пропуски зажигания.
Пропуски зажигания на ХХ или под давлением. Пропуски можно просканировать в каждом цилиндре при помощи ВАГ-КОМа.
Решение: Замена катушек зажигания или свечей. При пропусках возможно мигание ЧекЭнджина.____________________________________________________________________________________
7. Неисправность датчика давления топлива.
Симптомы:
Возможно появление ошибок?
012555 — Low Pressure Fuel regulation: Fuel Pressure Outside Specification
P310B — 008 — Implausible Signal — Intermittent — MIL ON
004767 — Low-Pressure Fuel Pressure Regulation: Pressure too High
P129F — 001 — Upper Limit Exceeded – Intermittent
— Машина глохнет при первом запуске «на холодную».
Решение: Замена датчика.
___________________________________________________________________________________
8. Цепной звук на холодную.
Симптомы:
Странный звук по утрам при запуске холодного двигателя. Звук появлялся при запуске и исчезал через 1-3 секунды.
Многие кидаются менять цепь рапредвалов. Не поможет, проверено 
Решение: Данный звук издаёт бендикс стартера.
___________________________________________________________________________________
9. Неисправность клапана рециркуляции N80 (EVAP)
Симптомы:
Увеличенный расход топлива и появился подсос воздуха. Когда сняли клапан — дулся во все стороны.
17834/P1426/005158 — продувочный клапан EVAP (N80): открытой цепи
Возможные причины:
-EVAP продувочный клапан (N80) неисправных
-Подключение / Разъемы / из EVAP продувочный клапан (N80) неисправных
-Реле топливного насоса (J17) неисправных
Возможные пути решения:
-Проверьте EVAP продувочный клапан (N80)
-Проверьте подключение / Разъемы / из EVAP продувочный клапан (N80)
-Проверка реле топливного насоса (J17)
а по VAGу так выдает :
ОШИБКИ:
1 неисправность:
17834 — EVAP Purge Valve (N80): Open Circuit
P1426 — 35-00 — —
Готовность: 0000 0000
Решение: Замена клапана. Номер 06E906517A
___________________________________________________________________________________
10. Залегание колец из-за странной их конструкции.
Симптомы: Увеличенный расход масла.
Решение: Лечится заменой маслосьемных колец с расширением канавки на поршне вверх, либо заменой целиком Шатунно-поршневой группы.
___________________________________________________________________________________
11. OIL PRESSURE на 55-65 градусов ож.
Симптомы: Загорается красная масленка пищит 3 раза OIL PRESSURE, далее уходит в поле музыкального трека, моргает там примерно от 3 до 10 раз и исчезает. Диагностикой ошибка не бьется в мозгах не прописывается и не фиксируется. Пищит именно в диапазоне 55-65 градусов ОЖ. Маслянный элемент при этом сильно мятый.
Решение: Замена масляного фильтра (элемента).
___________________________________________________________________________________
ИНФОРМАЦИЯ ВЗЯТА здесь и здесь и вот здесь . МНОЮ ДОПОЛНЕНЫ ПУНКТЫ 1 и 5 добавлен 11 .
Подскажите пожалуйста
1 неисправность:
0663 — Вакуумный датчик
C10A3 01 [137] — Сбой в электрике
[VAC_SENS_1_FAST_DATA_FAULT]
Индикатор неисправности ВКЛ — Подтверждено — протестировано после удаления записей
Стоп-кадр:
Биты состояния: 00000001
Приоритетность: 3
Счётчик возникновения ошибки: 3
Индекс забывания: 9
Код ошибки: 00 02 97
2779 — Датчик температуры охлаждающей жидкости(G62
P0116 00 [096] — )недостоверный сигнал
Непостоянно — Не подтверждено — протестировано после удаления записей
Стоп-кадр:
Биты состояния: 00000001
Приоритетность: 2
Счётчик возникновения ошибки: 1
Пробег: 62844 km
Дата: 2019.10.07
Время: 12:54:18
Engine speed: 0.00 /min
Нормированное значение нагрузки: 0.0 %
Скорость автомобиля: 0 km/h
Coolant temperature: 93 °C
Температура воздуха на впуске: 22 °C
Ambient air pressure: 980 mbar
Напряжение клеммы 30: 12.769 V
Индекс забывания по OBD: 40
Наружная температура: 2 °C
Абсолютное давление на впуске: 97 kPa abs
Engine torque: 0.0 Nm
Fuel level: 50.00 l
Лямбда-зонд: ряд 1: зонд 1: номинальное значение: 1.0278
Распредвал впускных клапанов: ряд 1: активный режим-Bits 0-7: 0
Распредвал впускных клапанов: ряд 1: активный режим-Bits 0-7: 0
Принудительный холостой ход-Bits 0-7: 0
10203 — Обнаружены пропуски воспламенения
P0300 00 [032] — —
Непостоянно — Не подтверждено — протестировано после удаления записей
Стоп-кадр:
Биты состояния: 00000001
Приоритетность: 2
Счётчик возникновения ошибки: 1
Пробег: 62783 km
Дата: 2019.10.05
Время: 20:12:23
Engine speed: 950.00 /min
Нормированное значение нагрузки: 21.6 %
Скорость автомобиля: 0 km/h
Coolant temperature: 51 °C
Температура воздуха на впуске: 10 °C
Ambient air pressure: 1000 mbar
Напряжение клеммы 30: 14.371 V
Индекс забывания по OBD: 36
Долговременная адаптация смесеобразования: ряд 1 на х.х.: -1.50 %
Лямбда-регулирование: адаптир. значение на част.нагрузке: ряд 1: 0.98633
Кратковременная адаптация смесеобразования: ряд 1: 0.0 %
Наружная температура: 8 °C
Проверка лямбда-зондов: ряд 1: разность лямбда: 0.00015
Fuel level: 52.00 l
10594 — Цил.4
P0304 00 [032] — обнаружены пропуски воспламенения
Непостоянно — Не подтверждено — протестировано после удаления записей
Стоп-кадр:
Биты состояния: 00000001
Приоритетность: 2
Счётчик возникновения ошибки: 1
Пробег: 62783 km
Дата: 2019.10.05
Время: 20:13:18
Engine speed: 950.00 /min
Нормированное значение нагрузки: 21.6 %
Скорость автомобиля: 0 km/h
Coolant temperature: 51 °C
Температура воздуха на впуске: 10 °C
Ambient air pressure: 1000 mbar
Напряжение клеммы 30: 14.371 V
Индекс забывания по OBD: 36
Наружная температура: 8 °C
Абсолютное давление на впуске: 29 kPa abs
Engine torque: 20.9 Nm
Fuel level: 52.00 l
Лямбда-зонд: ряд 1: зонд 1: номинальное значение: 1.0000
Распредвал впускных клапанов: ряд 1: активный режим-Bits 0-7: 0
Распредвал впускных клапанов: ряд 1: активный режим-Bits 0-7: 0
Принудительный холостой ход-Bits 0-7: 0
10592 — Цил.2
P0302 00 [032] — обнаружены пропуски воспламенения
Непостоянно — Не подтверждено — протестировано после удаления записей
Стоп-кадр:
Биты состояния: 00000001
Приоритетность: 2
Счётчик возникновения ошибки: 1
Пробег: 62783 km
Дата: 2019.10.05
Время: 20:17:32
Engine speed: 958.00 /min
Нормированное значение нагрузки: 25.1 %
Скорость автомобиля: 0 km/h
Coolant temperature: 66 °C
Температура воздуха на впуске: 12 °C
Ambient air pressure: 1000 mbar
Напряжение клеммы 30: 14.324 V
Индекс забывания по OBD: 36
Наружная температура: 9 °C
Абсолютное давление на впуске: 33 kPa abs
Engine torque: 24.2 Nm
Fuel level: 52.00 l
Лямбда-зонд: ряд 1: зонд 1: номинальное значение: 1.0000
Распредвал впускных клапанов: ряд 1: активный режим-Bits 0-7: 0
Распредвал впускных клапанов: ряд 1: активный режим-Bits 0-7: 1
Принудительный холостой ход-Bits 0-7: 0
что за ошибки и как их устронить
BrainyHacker
Опытный
Подскажите пожалуйста
1 неисправность:
0663 — Вакуумный датчик
C10A3 01 [137] — Сбой в электрике
[VAC_SENS_1_FAST_DATA_FAULT]
Индикатор неисправности ВКЛ — Подтверждено — протестировано после удаления записей
Стоп-кадр:
Биты состояния: 00000001
Приоритетность: 3
Счётчик возникновения ошибки: 3
Индекс забывания: 9
Код ошибки: 00 02 97
2779 — Датчик температуры охлаждающей жидкости(G62
P0116 00 [096] — )недостоверный сигнал
Непостоянно — Не подтверждено — протестировано после удаления записей
Стоп-кадр:
Биты состояния: 00000001
Приоритетность: 2
Счётчик возникновения ошибки: 1
Пробег: 62844 km
Дата: 2019.10.07
Время: 12:54:18
Engine speed: 0.00 /min
Нормированное значение нагрузки: 0.0 %
Скорость автомобиля: 0 km/h
Coolant temperature: 93 °C
Температура воздуха на впуске: 22 °C
Ambient air pressure: 980 mbar
Напряжение клеммы 30: 12.769 V
Индекс забывания по OBD: 40
Наружная температура: 2 °C
Абсолютное давление на впуске: 97 kPa abs
Engine torque: 0.0 Nm
Fuel level: 50.00 l
Лямбда-зонд: ряд 1: зонд 1: номинальное значение: 1.0278
Распредвал впускных клапанов: ряд 1: активный режим-Bits 0-7: 0
Распредвал впускных клапанов: ряд 1: активный режим-Bits 0-7: 0
Принудительный холостой ход-Bits 0-7: 0
10203 — Обнаружены пропуски воспламенения
P0300 00 [032] — —
Непостоянно — Не подтверждено — протестировано после удаления записей
Стоп-кадр:
Биты состояния: 00000001
Приоритетность: 2
Счётчик возникновения ошибки: 1
Пробег: 62783 km
Дата: 2019.10.05
Время: 20:12:23
Engine speed: 950.00 /min
Нормированное значение нагрузки: 21.6 %
Скорость автомобиля: 0 km/h
Coolant temperature: 51 °C
Температура воздуха на впуске: 10 °C
Ambient air pressure: 1000 mbar
Напряжение клеммы 30: 14.371 V
Индекс забывания по OBD: 36
Долговременная адаптация смесеобразования: ряд 1 на х.х.: -1.50 %
Лямбда-регулирование: адаптир. значение на част.нагрузке: ряд 1: 0.98633
Кратковременная адаптация смесеобразования: ряд 1: 0.0 %
Наружная температура: 8 °C
Проверка лямбда-зондов: ряд 1: разность лямбда: 0.00015
Fuel level: 52.00 l
10594 — Цил.4
P0304 00 [032] — обнаружены пропуски воспламенения
Непостоянно — Не подтверждено — протестировано после удаления записей
Стоп-кадр:
Биты состояния: 00000001
Приоритетность: 2
Счётчик возникновения ошибки: 1
Пробег: 62783 km
Дата: 2019.10.05
Время: 20:13:18
Engine speed: 950.00 /min
Нормированное значение нагрузки: 21.6 %
Скорость автомобиля: 0 km/h
Coolant temperature: 51 °C
Температура воздуха на впуске: 10 °C
Ambient air pressure: 1000 mbar
Напряжение клеммы 30: 14.371 V
Индекс забывания по OBD: 36
Наружная температура: 8 °C
Абсолютное давление на впуске: 29 kPa abs
Engine torque: 20.9 Nm
Fuel level: 52.00 l
Лямбда-зонд: ряд 1: зонд 1: номинальное значение: 1.0000
Распредвал впускных клапанов: ряд 1: активный режим-Bits 0-7: 0
Распредвал впускных клапанов: ряд 1: активный режим-Bits 0-7: 0
Принудительный холостой ход-Bits 0-7: 0
10592 — Цил.2
P0302 00 [032] — обнаружены пропуски воспламенения
Непостоянно — Не подтверждено — протестировано после удаления записей
Стоп-кадр:
Биты состояния: 00000001
Приоритетность: 2
Счётчик возникновения ошибки: 1
Пробег: 62783 km
Дата: 2019.10.05
Время: 20:17:32
Engine speed: 958.00 /min
Нормированное значение нагрузки: 25.1 %
Скорость автомобиля: 0 km/h
Coolant temperature: 66 °C
Температура воздуха на впуске: 12 °C
Ambient air pressure: 1000 mbar
Напряжение клеммы 30: 14.324 V
Индекс забывания по OBD: 36
Наружная температура: 9 °C
Абсолютное давление на впуске: 33 kPa abs
Engine torque: 24.2 Nm
Fuel level: 52.00 l
Лямбда-зонд: ряд 1: зонд 1: номинальное значение: 1.0000
Распредвал впускных клапанов: ряд 1: активный режим-Bits 0-7: 0
Распредвал впускных клапанов: ряд 1: активный режим-Bits 0-7: 1
Принудительный холостой ход-Bits 0-7: 0
что за ошибки и как их устронить
ChiPer
Админ
Подскажите пожалуйста
1 неисправность:
0663 — Вакуумный датчик
C10A3 01 [137] — Сбой в электрике
[VAC_SENS_1_FAST_DATA_FAULT]
Индикатор неисправности ВКЛ — Подтверждено — протестировано после удаления записей
Стоп-кадр:
Биты состояния: 00000001
Приоритетность: 3
Счётчик возникновения ошибки: 3
Индекс забывания: 9
Код ошибки: 00 02 97
2779 — Датчик температуры охлаждающей жидкости(G62
P0116 00 [096] — )недостоверный сигнал
Непостоянно — Не подтверждено — протестировано после удаления записей
Стоп-кадр:
Биты состояния: 00000001
Приоритетность: 2
Счётчик возникновения ошибки: 1
Пробег: 62844 km
Дата: 2019.10.07
Время: 12:54:18
Engine speed: 0.00 /min
Нормированное значение нагрузки: 0.0 %
Скорость автомобиля: 0 km/h
Coolant temperature: 93 °C
Температура воздуха на впуске: 22 °C
Ambient air pressure: 980 mbar
Напряжение клеммы 30: 12.769 V
Индекс забывания по OBD: 40
Наружная температура: 2 °C
Абсолютное давление на впуске: 97 kPa abs
Engine torque: 0.0 Nm
Fuel level: 50.00 l
Лямбда-зонд: ряд 1: зонд 1: номинальное значение: 1.0278
Распредвал впускных клапанов: ряд 1: активный режим-Bits 0-7: 0
Распредвал впускных клапанов: ряд 1: активный режим-Bits 0-7: 0
Принудительный холостой ход-Bits 0-7: 0
10203 — Обнаружены пропуски воспламенения
P0300 00 [032] — —
Непостоянно — Не подтверждено — протестировано после удаления записей
Стоп-кадр:
Биты состояния: 00000001
Приоритетность: 2
Счётчик возникновения ошибки: 1
Пробег: 62783 km
Дата: 2019.10.05
Время: 20:12:23
Engine speed: 950.00 /min
Нормированное значение нагрузки: 21.6 %
Скорость автомобиля: 0 km/h
Coolant temperature: 51 °C
Температура воздуха на впуске: 10 °C
Ambient air pressure: 1000 mbar
Напряжение клеммы 30: 14.371 V
Индекс забывания по OBD: 36
Долговременная адаптация смесеобразования: ряд 1 на х.х.: -1.50 %
Лямбда-регулирование: адаптир. значение на част.нагрузке: ряд 1: 0.98633
Кратковременная адаптация смесеобразования: ряд 1: 0.0 %
Наружная температура: 8 °C
Проверка лямбда-зондов: ряд 1: разность лямбда: 0.00015
Fuel level: 52.00 l
10594 — Цил.4
P0304 00 [032] — обнаружены пропуски воспламенения
Непостоянно — Не подтверждено — протестировано после удаления записей
Стоп-кадр:
Биты состояния: 00000001
Приоритетность: 2
Счётчик возникновения ошибки: 1
Пробег: 62783 km
Дата: 2019.10.05
Время: 20:13:18
Engine speed: 950.00 /min
Нормированное значение нагрузки: 21.6 %
Скорость автомобиля: 0 km/h
Coolant temperature: 51 °C
Температура воздуха на впуске: 10 °C
Ambient air pressure: 1000 mbar
Напряжение клеммы 30: 14.371 V
Индекс забывания по OBD: 36
Наружная температура: 8 °C
Абсолютное давление на впуске: 29 kPa abs
Engine torque: 20.9 Nm
Fuel level: 52.00 l
Лямбда-зонд: ряд 1: зонд 1: номинальное значение: 1.0000
Распредвал впускных клапанов: ряд 1: активный режим-Bits 0-7: 0
Распредвал впускных клапанов: ряд 1: активный режим-Bits 0-7: 0
Принудительный холостой ход-Bits 0-7: 0
10592 — Цил.2
P0302 00 [032] — обнаружены пропуски воспламенения
Непостоянно — Не подтверждено — протестировано после удаления записей
Стоп-кадр:
Биты состояния: 00000001
Приоритетность: 2
Счётчик возникновения ошибки: 1
Пробег: 62783 km
Дата: 2019.10.05
Время: 20:17:32
Engine speed: 958.00 /min
Нормированное значение нагрузки: 25.1 %
Скорость автомобиля: 0 km/h
Coolant temperature: 66 °C
Температура воздуха на впуске: 12 °C
Ambient air pressure: 1000 mbar
Напряжение клеммы 30: 14.324 V
Индекс забывания по OBD: 36
Наружная температура: 9 °C
Абсолютное давление на впуске: 33 kPa abs
Engine torque: 24.2 Nm
Fuel level: 52.00 l
Лямбда-зонд: ряд 1: зонд 1: номинальное значение: 1.0000
Распредвал впускных клапанов: ряд 1: активный режим-Bits 0-7: 0
Распредвал впускных клапанов: ряд 1: активный режим-Bits 0-7: 1
Принудительный холостой ход-Bits 0-7: 0
что за ошибки и как их устронить
Проводится активный набор модераторов, подробности здесь.
Прежде чем задать на форуме вопрос — убедитесь, что вы будете первым.
Внимательно ознакомьтесь с Правилами форума .
Поддержать проект:
Яндекс Деньги: 410012123136543
BTC: 1C2sJoVR4w2PQ1kxYsE4NccXv2ZLFMqT3G

Автор: Валерия Филинович 24.07.2020 Время прочтения: 4 минут 26 268 просмотров Комментариев пока нет
Конструктивно самолет не сложнее современной легковушки. Но в кабине пилота, бесчисленное количество кнопок и тумблеров, а на торпеде автомобиля их нет. Почему так?
Причин несколько. Во-первых, эстетика и комфорт. Во-вторых, пилоту жизненно важно диагностировать проблему сразу. «На земле» у автомобилиста всегда найдется время заехать на диагностику. В-третьих, дилеры хорошо зарабатывают на диагностике.
Поэтому автопроизводители предусматривают систему контрольных ламп на приборном щитке, которые сигнализируют о проблемах в технике или электронике. Одним из таких индикаторов служит значок EPC. Что он значит? Чего бояться и куда бежать? Об этом ниже.
Что означает значок ЕРС на приборной панели Ауди
Чаще всего «горит» EPC на приборной панели Ауди, VW и Porsche. Концерн VAG использует одни и те же электронные «мозги». С небольшими отличиями, разумеется.
Что касается EPC, аббревиатура расшифровывается как «электронное управление мощностью». Многие автомобилисты отмечают, что, когда загорается EPC, Audi просто «не едет», урезаны обороты, троит двигатель (если с электронным впрыском) и т.д.
Официальный комментарий Ингольштадта звучит так: «EPC Ауди — это система информирования о неисправности электронных систем автомобиля. До устранения причин сигнала, электроника временно ограничивает подачу топлива и, при необходимости, отключает ряд бортовых систем». То есть, если на Ауди горит EPC, значит что-то не так с электроникой и для точного выяснения причин нужно пройти диагностику в сервисном центре.
Обратите внимание! У многих немецких автомобилях при повороте ключа зажигания индикатор EPC загорается на пару секунд, после чего гаснет. Это значит, что система работает нормально, а вот если горит постоянно — это повод обратиться в сервис.
Ошибка EPC на Audi A3
В каких случаях он загорается
Индикатор EPC загорается при обнаружении любой проблемы, связанной с бортовой электроникой и реакцией датчиков на неисправности механического характера. Зачастую, контрольная лампа свидетельствует о:
- неисправности всевозможных датчиков (давления и герметичности впускного коллектора, охлаждения, температуры, круиза, климата, света, дождя, колена и пр.);
- проблемах с байпасом;
- неисправности блока дроссельной заслонки;
- наличии «пропусков» в цилиндрах;
- сгорании клапана;
- проблемах с вращением коленвала;
- отказе сервопривода дросселя;
- низком/высоком сигнале педали газа или тормоза;
- неисправности систем пассивной безопасности и т.д.
Определить проблему на глаз не получится. При подключении компьютера или смартфона система сама покажет номер ошибки. После чего остается посмотреть, что она значит.
Бывают и не стандартные ситуации. Например, EPC ошибка на Ауди показывает неисправность датчика давления в картере. Но на деле с ним все в порядке. Оказалось, что была плохо закручена крышка горловины после долива масла (нужно до щелчка). В системе шел подсос воздуха и давление в картере оказалось ниже нормы, что привело к срабатыванию контрольной лампы. Но это исключительный случай.
Безопасно ли ездить с таким индикатором
Учитывая, что контрольная лампа может гореть даже от банального перегорания лампочки или реле, то в каких-то случаях можно ездить и не беспокоиться. Например, неисправный датчик дождя для многих не станет весомым поводом обращаться в сервис и платить от 1000 руб. за диагностику.
Другое дело, если вместе с EPC загорается лампа «Check Engine» и отсутствует «драйв» двигателя. В этом случае тянуть с диагностикой нельзя. В противном случае в ближайшие 100 км не избежать серьезных механических повреждений (замены поршневой, ГРМ и пр.).
Ошибки EPC по моделям
Ошибок и неисправностей, о которых сигнализирует EPC — сотни. Всех, естественно, не перечислить. Все рассчитано на то, чтобы автовладелец обратился в сервисный центр. Но, как говорится, у каждой модели есть свои характерные проблемы, на которые жалуются владельцы на профильных форумах. Ниже краткая выжимка по некоторым моделям бренда.
Индикатор EPC на приборной панели у владельцев Ауди Q5 часто выдает следующие ошибки:
- Ошибка сопровождается индикатором «Check Engine» и говорит о неисправности процессора блока управления КПП. Потребуется, либо замена блока целиком, либо разъема, либо нескольких проводов;
- Данная ошибка EPC на Audi Q5 свидетельствует о повреждении концевого выключателя идущего на педаль сцепления. Только замена;
- Ошибка высвечивается при неисправности реле муфты компрессора климата. Ремонта заключается в замене реле или самой муфты;
- Сбой модуля управления свечами. Требуется замена проводов, свечей, реле или самого модуля.
Что интересно, у многих владельцев Audi Q5 EPS загорался, когда Ингольштадт выкатывал новую прошивку, которую предлагалось установить в добровольно-принудительном порядке.
Ошибка EPC на приборке Audi Q5
Если посмотреть «ветку» владельцев модели A4, тут выявляются свои «болячки». Например:
- Отсутствие сигнала от турбины гидротрансформатора АКПП. Проблема решается заменой масла, датчика температуры или корпуса клапана. По ситуации;
- Ошибочный сигнал от датчика оборотов двигателя. Потребуется его замена или визуальный осмотр на наличие коррозийных проводов;
- Проблема с передаточным отношением в КПП. Ремонт зависит от характера поломки. Начиная от замены проводов и масляного насоса, заканчивая полной заменой коробки.
Также, конкретно на Ауди А4 B8 ошибка P039в в EPC свидетельствует о высоком давлении в первом цилиндре. Возможна проблема с турбокомпрессором. Распространенная болячка.
Как убрать ошибку УЗС на Ауди А4 1.8Т, смотрите в видеоролике ниже:
По Ауди А6 автомобилисты также часто спрашивают по таким ошибкам, как:
- Неисправность в датчике скорости. Ремонт может не ограничится парой проводов или самим датчиком. Периодически приходится менять весь модуль ECM;
- Высокая температура модуля управления мотором. При ремонте проверяется заземление и предохранители. При необходимости модуль меняется;
- Ненормированное напряжение во входной цепи уровня топлива. Ремонт может ограничится заменой проводов и соединителей, либо заменой модуля РСМ.
Также, если брать конкретно Ауди A6 C5, в EPC частая ошибка P1676, которая свидетельствует об обрыве цепи контрольной лампы левого стоп-сигнала. Ремонтируется путем поиска обрыва, проверки изоляции и замены коррозийных проводов.
Резюмируя, ошибок в системе EPC сотни и каждая сигнализирует об определенной проблеме. Это значительно упрощает ремонт высокотехнологичных авто и помогает вовремя предупредить серьезные неисправности двигателя и КПП, пока стоимость ремонта не превысила стоимость самой машины.

Сеть магазинов «ВкусВилл» удалила ставшую причиной резкой критики статью, в которой в числе героев были четыре женщины, члены одной семьи, из них две девушки обручены. Компания извинилась перед теми, чьи чувства задела публикация.
«На этом месте была статья, которая больно задела чувства большого числа наших покупателей, сотрудников, партнеров и поставщиков. Мы сожалеем, что так получилось, и считаем эту публикацию своей ошибкой, ставшей проявлением непрофессионализма отдельных сотрудников», — говорится в сообщении компании.
Цель компании — дать возможность людям покупать продукты, а не «публиковать статьи, которые являются отражением каких-либо политических или социальных взглядов», подчеркнули в сети.
Обращение с извинениями подписали руководители компании, в том числе ее основатель Андрей Кривенко, управляющий информацией Валерий Разгуляев, управляющая единой концепцией Алена Несифорова и другие.
Одна из героинь статьи рассказала РБК, что сеть не связывалась с ними по поводу удаления материала. «ВкусВилл» с нами не связывался, ничего сказать по этому поводу я не могу», — пояснила она. «С выхода текста нам начали поступать угрозы во все аккаунты моей семьи», — добавила она.

Материал был опубликован в разделе «Школа потребителя», где сеть рассказывает о своих клиентах. Героями публикации были разные семьи: без детей, многодетные, с домашними животными, а также семья из четырех женщин. Членами семьи являются Юма, а также две ее дочери, одна из которых недавно обручилась с девушкой. В материале была опубликована их фотография.
Герои статьи рассказывали, какие продукты они покупают в магазинах сети, кто в их семье готовит, кто ходит за продуктами. В статье также была пометка о том, что ей присвоена категория «18+» по требованию российского законодательства, так как одна из девушек — несовершеннолетняя.
По словам Полякова, его подразделение не обязано согласовывать каждую публикацию с руководством и о том, что на сайте выйдет фотография женщин, оно узнало уже после публикации.
После публикации на «ВкусВилл», а также на девушек из материала в соцсетях обрушилась резкая критика. Комментарии появились в соцсетях «ВкусВила», а также в аккаунтах героинь.
Предлагаем расшифровку некоторых выявленных бортовым компьютером нарушений.

Общие принципы
После включения зажигания, электронная система управления проверяет все узлы и устройства автомобиля. Обнаруженные проблемы делятся на первостепенные и обозначаются красными пиктограммами, и второстепенные: обозначены желтыми пиктограммами.
При нарушениях первостепенной важности раздается три звуковых сигнала. Если нарушений несколько, разные пиктограммы появляются, сменяя друг друга каждые 2 секунды.
Нарушения второстепенной важности сопровождаются однократным звуковым сигналом.
Расшифровка сообщений
SERVISE — так режимный сервисный индикатор напоминает о плановом ТО. Надпись появляется за 3 000 км до срока ТО. Когда он наступает, владелец видит на приборной панели надпись SERVISE JETZT
AIRBAG FEHLER! — эта надпись говорит о неисправности в системе подушек безопасности. В норме, после включения зажигания несколько секунд горит контрольный индикатор, затем он должен погаснуть. Если подушки отключены, лампа мигает 12 секунд. Если лампочка загорается во время движения автомобиля, и вы видите надпись — система подушек неисправна
BEIM EINLEGEN EINER FAHRSTUFF IM STAND FUSSBREMSE BETATIGEN! — такая надпись появляется на автомобилях с автоматической коробкой передач. Это напоминание: перед выбором позиции селектора АКПП выжмите педаль тормоза
BREMSBELAG PRUFEN — надпись указывает на износ тормозных колодок и переводится как «Проверьте состояние тормозных колодок»
STOP! BREMSENFEHLER BETRIEBSANLEITUNG! — очень важная надпись. «Остановись! Тормозная система неисправна!». Лампочка ABS загорается в норме после включения зажигания, затем гаснет. Если она не загорелась вообще, или не погасла после запуска, или загорелась в движении — система ABS не в порядке. Если горит и ABS, и лампа тормозов — продолжать движение нельзя!
Еще STOP! BREMSENFEHLER BETRIEBSANLEITUNG может загореться, если уровень тормозной жидкости в системе упал
GENERATOR WERKSTATT! — переводится как «Батарея разряжена». В норме, сигнальная лампа заряда аккумулятора гаснет после запуска мотора. Продолжает гореть — значит, АКБ разряжается
MOTORSTORG WERKSTATT! — нарушения в работе двигателя. Переводится как «неполадки в двигателе». Часто сочетается с лампочкой EPC — сигнальной лампы системы управления двигателем. Продолжать движение нельзя
STOP! KUHLMITTEL PRUFEN BETRIBSANLEITUNG! — упал уровень антифриза в системе охлаждения двигателя. Переводится как «Остановись! Проверь охлаждающую жидкость»
STOP! OLDRUCK MOTOR AUS! — упало давление в системе смазывания двигателя. Переводится как «Остановить! Нет давления масла!». Если сопровождается горящим красным индикатором «масленки» и троекратным звуковым сигналом — движение продолжать нельзя
STOP! OLSTAND PRUFEN! — сигнал о том, что необходимо проверить уровень масла в двигателе. Сочетается с желтой «масленкой» на панели приборов. Ехать можно, но нужно долить масло в двигатель по щупу.
OLSENSOR WERKSTATT! — «Неисправен датчик уровня масла». Встречается вместе с мигающей желтым цветом «масленкой»
WASCHWASSER AUFFULLEN — сообщение о том, что жидкость омывателя в резервуаре заканчивается, и нужно ее долить
BITTE TANKEN — «Пожалуйста, заправьтесь»
Другие сообщения бортовой системы контроля
- Siehe Betriebsanleitung — Смотрите инструкцию
- Parkbremse losen- Отпустите стояночный тормоз
- Bremstlussigkeit prufen — Провеьте уровень тормозной жидкости в системе
- Kullwassertemperatur — высокая температура охлаждающей жидкости
- ASC — Включен автоконтролер стабилизации движения
- Bremslichtelektrik — световой сигнал торможения не работает
- Niveauregelung — Низкий уровень подкачки задних амортизаторов
- Kofferraum offen- Багажник открыт
- Tur-off — дверь открыта
- Check-Control — неисправность в контроллере ошибок (бортовой компьютер вышел из строя)
- Getribenotprogramm — отказ управления коробки передач
- Funkschlussel Batterie — сели батарейки ключа дистанционного управления
- Bremsdruck — низкое давление в тормозной системе
Prufen von — проверьте
- Bremslicht — сигналы торможения
- Abblendlicht — ближний свет
- Standlicht — стояночные огни
- Rucklicht — свет задних фонарей
- Nebellicht vorne / hinten — противотуманный свет спереди / сзади
- Kennzeichenlicht — освещение номерного знака
- Anhangerlicht — фонари прицепа
- Fernlicht — дальний свет
- Ruckfabrlicht- свет заднего хода
- Getriebe — электрику АКПП
- Sensor-Oilstand — датчик уровня моторного масла
- Oilstand Getriebe — уровень масла в АКПП
Еще больше расшифровок ошибок на приборной панели автомобилей VAG:
Читайте также:
- Ошибка 1550 опель зафира
- P0341 ошибка ssangyong kyron
- P1121 ошибка маз 651
- Ошибка b109d71 шкода октавия а7
- Ошибка 2148 фиат дукато