

12. 09. 2017 Тема урока: Мутации

Термин «мутация» впервые предложил голландский ботаник Гуго де Фриз в 1901 году. Он к выводу, что вид может распадаться на различные виды, наблюдая изменчивость ослинника. Это явление Де Фриз назвал мутациями, считая что биологические виды время от времени находятся в фазе мутирования. Разработал мутационную теорию. В 1877 году впервые измерил осмотическое давление у растений, ввёл понятия плазмолиз и деплазмолиз. Переоткрыл и подтвердил в 1900 году, одновременно с К. Э. Корренсом и Э. Чермак Зейзенеггом законы Грегора Менделя

Ошибка

• Жизнь — это задача, которую, еще ни кто не РЕШИЛ БЕЗ ОШИБОК. • Некоторые ошибки в жизни являются опечатками, которые впоследствии впечатываются в память навсегда.

Польза Ошибка Вред

• Мутации это новообразования, возникающие в результате ошибок деления клетки. • Мутациями называются внезапные изменения генотипа, влекущие изменения признаков, которые наследуются потомством.

Значение мутаций в эволюции • Мутации являются материалом для естественного отбора.

Виды мутаций • Генные –изменения в пределах одного гена • Хромосомные – изменения строения хромосом • Геномные – изменения количества хромосом
 числа геномов Генные Изменение числа отдельных хромосом Хромосомные Внутрихромосомные Межхромосомные")
Мутации Геномные Увеличение (уменьшение) числа геномов Генные Изменение числа отдельных хромосом Хромосомные Внутрихромосомные Межхромосомные Полиплоидия Гаплоидия Триплоидия Дупликация, амплификация Дефишенси «Центрическое разделение»

Хромосомные мутации Дупликация — удвоение участка хромосомы

Хромосомные мутации Инверсия поворот участка на 1800 градусов

Хромосомные мутации Транслокация — перемещение участка одной хромосомы на другую, негомологичную хромосому

Хромосомные мутации Дефишенси
 мутации • Дупликация. В этом случае происходит повторное дублирование либо удвоение фрагмента")
Генные (точковые) мутации • Дупликация. В этом случае происходит повторное дублирование либо удвоение фрагмента ДНК. • Делеция. В этом случае имеет место утрата фрагмента ДНК. • Инверсия. В этом случае отмечается поворот на 180 град. участка ДНК. Его размер может быть как в два нуклеотида. • Инсерция. В этом случае происходит вставка участков ДНК.
 мутации")
Генные (точковые) мутации

Кратное увеличение набора хромосом Полиплоидия Геномные Изменение количества Гаплоидия хромосом Триплоидия Утеря или проявление лишних хромосом Анеуплоидия 2 n+1 в результате нарушения мейоз Гетероплоидия Дупликация, амплификация Удвоеный участок может сохранять свое положение, Изменение поворачиваться на 180°, или даже повторяться несколько Хромосомные структуры раз, и тогда такая мутация называется амплификацией. хромосом. АБВГДВГДЕЖЗИК Потеря (выпадение) участка хромосомы АБЕЖЗИК Делеция Нормальн Поворот участка на 1800 градусов АБДГВЕЖЗИК Инверсия ый Перемещение участка одной хромосомы на другую, негомологичную АБВГЛМНОПДЖЗ Транслокация Концевые нехватки ВГДЕЖЗИК Межхромос омные порядок генов АБВГДЕЖЗ ИК Внутрихромосомные Изменение Дефишенси Слияние негомологичных хромосом в одну хромосому. Центрическое слияние Разделение хромосом на две. Центрическое разделение Выпадение одного или нескольких нуклеотидов из молекулы

Проверь себя а б в г д е
 является следствием ошибок при репликации ДНК; б)")
Проверь себя Характеристики мутаций Виды мутаций а) является следствием ошибок при репликации ДНК; б) приводит к увеличению числа хромосом; в) приводит к образованию новой формы гена; г) изменяют последовательность генов в хромосоме; д) наблюдается у растений; е) затрагивают отдельные хромосомы. А) геномные; Б) хромосомные; В) генные.

Проверь себя а б в г д е В А В Б А Б

Классификация мутаций

По характеру изменения генотипа • Генные –изменения в пределах одного гена • Хромосомные – изменения строения хромосом • Геномные – изменения количества хромосом

По причины мутаций • Мутагены – факторы, вызывающие мутации: физические – например, температура, радиация химические – например, НNO 2, иприт биологические например, вирусы

По адаптивному значению

Не все мутации приводят к изменениям генотипа Аланин Глицин – Лизин – Изолейцин ЦГА — ЦЦА — ТТЦ — ТАА Аланин Глицин – Лизин – Изолейцин ЦГА ЦЦТ ТТЦ ТАА

По месту их возникновения

По месту характеру проявления

По проявлению в фенотипе Биохимические Физиологические Морфологические

• Свойства мутаций • Мутации явление редкое. • По своему проявлению мутации могут быть полезными и вредными, доминантными и рецессивными. • Чаще бывают рецессивные мутации, чем доминантные. Рецессивные мутации накапливаются в генофонде длительно не проявляя себя. • Мутации возникают внезапно, скачкообразно. • Мутации наследуются, т. е. стойко передаются из поколения в поколение. • Мутации могут возникать повторно.

• Свойства мутаций • Мутации ненаправленные мутировать может любой ген, вызывая изменения как незначительных, так и жизненно важных признаков. • Способность к мутировать— одно из свойств гена. • Каждая отдельная мутация вызывается какой то причиной, но в большинстве случаев эти причины неизвестны. • Мутации связаны с изменениями во внешней среде. • Это убедительно доказывается тем, что путем воздействия внешними факторами удается резко повысить их число.

• Что дают мутации эволюционному процессу? • Природа идет по пути усложнения своей организации • Сложные биологические системы, организмы – открывают новые возможности для проявления жизни. • Что видит дождевой червь и что видит человек своим глазным яблоком? • Внезапны, скачкообразны, их нельзя «вычислить» …. Тем не менее, они целесообразны. И эта целесообразность возникает из число малых нецелесообразных проб. • Мутации являются материалом для естественного отбора.

Факторы вызывающие мутации • Мутагены бывают: • Естественные и искусственные • Экзогенные и эндогенные

По природе возникновения мутагены классифицируют Физические: Биологические: * некоторые вирусы * ионизирующее излучение; (вирус кори, краснухи, гриппа); * радиоактивный распад; * ультрафиолетовое излучение; * антигены некоторых микроорганизмов; * продукты обмена веществ (продукты * моделированное окисления липидов). радиоизлучение и электромагнитные поля; * чрезмерно высокая или низкая температура.
; * алкилирующие агенты")
Химические мутагены * окислители и восстановители (нитраты, нитриты, активные формы кислорода); * алкилирующие агенты (например, иодацетамид); * пестициды (например гербициды, фунгициды); * некоторые пищевые добавки (например, ароматические углеводороды, цикламаты); * продукты переработки нефти; * органические растворители; * лекарственные препараты (например, цитостатики, препараты ртути, иммунодепрессанты).

Сравнительная характеристика форм изменчивости Характеристика Модификационная изменчивость Мутационная изменчивость Объект изменения Отбирающий фактор Наследование Значение для особи Фенотип в пределах нормы реакции Генотип Изменение условий окружающей среды Не наследуются Понижается или повышается жизнеспособность Способствует выживанию Некоторые условий окружающей среды Роль в эволюции Форма изменчивости Подчиненность закономерности Приспособление организма к условиям Материал для естественного отбора среды Определенная (групповая) Неопределенная (индивидуальная) Статистическая закономерность вариационных рядов Наследуются Полезные приводят к победе в борьбе за существование, вредные – к гибели. Приводит к образованию новых популяций, видов Закон гомологических рядов наследственной изменчивости


Использование мутагенеза

• Мутагенез широко применяется в селекции микроорганизмов и растений.
 Гексоплоидное растение (6 n)")
полиплоидия Диплоидное растение (2 n) Гексоплоидное растение (6 n)

Использование полиплоидов человеком

Наследственные болезни человека

 мутации")
Генные (Точковые) мутации

Генная мутация Гемофилия наследование по Х-хромосоме, рецессивное • Гемофилия — наследственное заболевание, связанное с нарушением коагуляции (процесс свёртывания крови); при этом заболевании возникают кровоизлияния в суставы, мышцы и внутренние органы, как спонтанные, так и в результате травмы или хирургического вмешательства. • Обычно гемофилией болеют мужчины, а женщины являются носителем больного гена.

Гемофилия

Генная мутация Дальтонизм Наследование по Х-хромосоме, рецессивное Дальтони зм, цветовая слепота — наследственная, реже приобретённая особенность зрения, выражающаяся в неспособности различать один или несколько цветов. Названа в честь Джона Дальтона, который впервые описал один из видов цветовой слепоты на основании собственных ощущений, в 1794 году.

Генная мутация Альбинизм • Альбинизм — врождённое отсутствие пигмента кожи, волос, радужной и пигментной оболочек глаза. Внешние проявления Могут наблюдаться изменения в сетчатке, возникать различные расстройства зрения, в том числе близорукость, дальнозоркость и астигматизм, а также повышенная чувствительность к свету и другие аномалии. Люди-альбиносы имеют белую окраску кожи; волосы у них белые (или они блондины). Частота альбиносов у народностей европейских стран оценивается примерно как 1 на 20 000 жителей. У некоторых других народностей альбиносы встречаются чаще. «Белая ворона»

Генная мутация Ихтиоз • Выраженность болезни проявляется диффузными нарушениями ороговения кожи – кератинизации, процессу, в результате которого происходит обезвоживание или гибель эпидермальных клеток с отторжением верхних слоев кожи и образованием плотных чешуек. • Нарушена температурная регуляция кожи, водный баланс, рост и развитие. • При жестоких формах ихтиоза младенцы умирают вскоре после рождения от обезвоживания и инфекций. • Задержка умственного развития, иммунодефицит, глухота, облысение, деформация костей, плохое зрение, психологическая опустошённость.
. • Практически")
• Генная мутация • Нейрофиброматоз • Тип наследования АД (аутосомно доминантное). • Практически в половине случаев болезнь Реклингхаузена возникает из за мутации в 17 -й и 22 -й хромосомах. • Для заболевания характерно появление множественных пигментированных пятен цвета «кофе с молоком» , доброкачественных новообразований — нейрофибром, опухолей центральной нервной системы, костных аномалий, изменений радужной оболочки глаза и целого ряда других симптомов.

• Генная мутация • в 7 -ой хромосоме

• Генная мутация • Фенилкетонурия

Генная мутация Серповидно-клеточная анемия. В состав цепи гемоглобина входит 146 аминокислотных остатков, которые закодированы в ДНК в виде 146 триплетов (438 нуклеотидов). ГАА – ГЛУТАМИНОВАЯ КИСЛОТА ГТА — ВАЛИН

Генная мутация Серповидно-клеточная анемия. Дефект гена в результате замены валина на глутаминовую кислоту. Серповидные эритроциты вызывают увеличение вязкости крови, создают механическую преграду в мелких артериолах и капиллярах, приводя к тканевой ишемии (с чем связаны болевые кризы). Кроме того, серповидные эритроциты менее устойчивы к механическим воздействиям, что приводит к их разрушению. Клиническая картина: умеренная желтуха, трофические язвы в области лодыжек, отставание в физическом развитии (особенно у мальчиков). ГАА – ГЛУТАМИНОВАЯ КИСЛОТА ГТА ВАЛИН

• Генная мутация • Миодистрофия Дюшена • Болезнь вызывается делециями или дупликациями одного или нескольких экзонов, либо точечными мутациями в гене дистрофина. (тип наследования Х-сцепленный рецесивный). Болеют мальчики и очень редко девочки. Основное проявление — слабость мышц, затруднения при движениях с детского возраста, которые прогрессируют с течением времени. Признаками этого заболевания являются специфическая походка и осанка страдающих им мальчиков, позднее начало ходьбы, ухудшенная по сравнению со сверстниками речь.
 мутация Синдром Тричера Коллинза Он обнаруживается у 1 из 50 тысяч детей.")
Точковая (Генная) мутация Синдром Тричера Коллинза Он обнаруживается у 1 из 50 тысяч детей. Характерна деформацией черепно лицевых костей. Причина дефект в гене TCOF 1, расположенном на q-плече 5 хромосомы.

• Генная мутация. Болезнь бабочки • Заболевание является результатом мутаций в более чем 10 генах • Буллёзный эпидермолиз (БЭ) — группа генетически и клинически гетерогенных заболеваний, характеризующаяся образованием пузырей и эрозий на коже и слизистых оболочках, ранимостью кожи и её чувствительностью к незначительной механической травме. • Наследуется БЭ как по аутосомнодоминантному, так аутосомно-рецессивному типу. • Частота встречаемости различных типов БЭ варьирует от 1: 30 000 до 1: 1 000 и также зависит от популяции.
 • «Гуттаперчевый мальчик» Д. В. Григорович •")
• Синдром дисплазии соединительной ткани (СДСТ) • «Гуттаперчевый мальчик» Д. В. Григорович • Единственной системой, способной как то компенсировать недостаточность соединительной ткани, является мышечная система. Отсюда и образ жизни гуттаперчевых детей. У этих детей должна быть хорошо развита мышечная система, и не только мышцы скелета, но и мышцы глаз, сердца и др. органов

Генная мутация Синдром Марфана • Дифференцированная форма врожденной соединительнотканной недостаточности, характеризующаяся разнообразными проявлениями скелетной, сердечно сосудистой и глазной патологии. У больных с синдромом Марфана отмечаются гигантизм, долихостеномелия и арахнодактилия, аневризмы аорты, миопия, эктопия хрусталика, деформация грудины, кифосколиоз, плоскостопие
 это группа наследственных")
• Синдром Элерса-Данло (АД, АР и Х сцепленный типы наследования) это группа наследственных системных заболеваний соединительной ткани, вызванных дефектом в синтезе коллагена.

• Синдром Стиклера. Наследуется АД • Характеризующееся прогрессирующей миопией, деградацией стекловидного тела, отслойкой сетчатки, прогрессирующей нейросенсорной тугоухостью, расщелиной твердого нёба, гипоплазией нижней челюсти, гипер и гипо мобильностью суставов, дисплазией эпифизов и потерей трудоспособности из за боли в суставах, вывихов и дегенеративных изменений.

Хромосомные мутации

Хромосомная мутация Концевая нехватка Делеция 5 -ой хромосомы Синдром кошачьего крика • Характерно: нарушение функций сердечнососудистой, пищеварительной систем, недоразвитие гортани (с характерным криком, напоминающим кошачье мяуканье), общее отставание развития, умственная отсталость, лунообразное лицо с широко расставленными глазами, деформированные низко расположенные ушные раковины. • Синдром встречается у 1 новорожденного из 50 000.
 Синдром Вильямса • Потеря участка (делеция) 7 -ой хромосомы. В •")
Потеря участка (делеция) Синдром Вильямса • Потеря участка (делеция) 7 -ой хромосомы. В • • • утраченном так называемом «длинном плече хромосомы» содержится около 26 генов. Больные имеют особое строение лица, в специальной литературе называемое «лицом эльфа» . у них низко расположенные глаза, окруженные припухлыми веками (что особенно заметно, когда человек смеется); большой рот с пухлыми губами и неправильным прикусом; очень широкий лоб; нос короткий, с плоской переносицей и круглым тупым кончиком; подбородок слегка заостренный; низко посаженные уши; выпуклый затылок; ярко голубые глаза (глазные яблоки тоже имеют голубоватый оттенок)
 Синдром Вильямса • Степень умственного развития детей с синдромом «лица эльфа»")
Потеря участка (делеция) Синдром Вильямса • Степень умственного развития детей с синдромом «лица эльфа» значительно варьирует – от легкой степени до значительного интеллектуального дефекта. • Для таких больных характерно отсутствие способности к наглядно образному мышлению, концентрированию на поставленной задаче, планированию и организации деятельности. • Люди с синдромом Вильямса отличаются коммуникабельностью, добродушием. • Но уже даже будучи взрослыми, они проявляют эмоциональную лабильность, излишнюю непосредственность и инфантильность, неадекватность в поведении, им свойственны различные фобии, навязчивые действия.

Геномные мутации

Генная мутация ХХY Синдром Клайнфельтера Встречается у 1 из 500 мальчиков.

Генная мутация ХО Синдром Шерешевского — Тернера

• Синдром Эдвардса Трисомии 18 хромосомы • Для него характерны множественные пороки развития, наиболее частыми являются задержка внутриутробного развития, ВПС, низко посаженные аномальной формы ушные раковины, короткая шея. Синдром имеет неблагоприятный прогноз, поэтому кардиохирурги не берут таких детей на хирургическую коррекцию порока сердца.
 Резкая умственная отсталость")
Синдром Патау Трисомии 13 хромосомы • • Микроцефалия (уменьшение головного мозга) Резкая умственная отсталость Расщепление верхней губы и неба Аномалии глазного яблока Повышенная гибкость суставов Полидактилия Высокая смертность (в первый год жизни умирает 90% детей)

Трисомия 21 -й хромосомы

• В мире в год появляется на свет 2 миллиона таких детей. • Среди них — 150 тысяч с синдромом Дауна

• 1. Выберите два верных ответа из пяти. • Искусственный мутагенез используют для • 1) получения организмов с новыми свойствами; • 2) сохранения полезных свойств организма; • 3) получения новых пород млекопитающих животных; • 4) выведения чистых линий; • 5) получения новых штаммов бактерий.

2. Выберите два верных ответа из пяти. • Мутации со сходным фенотипическим проявлением могут появиться, скорее всего, у овса и • 1) подсолнуха; • 2) ржи; • 3) картофеля; • 4) гороха; • 5) риса.

Задание № 1 • Представьте, что вы селекционеры, вам необходимо создать новый сорт томатов, которые будут выращиваться для приготовления томатного сока? Какими признаками бы вы наделили свой сорт? Почему? Задание № 2 • Вам необходимо вывести новый сорт томата, пригодного для механизированной уборки плодов. Какими признаками вы наделили бы свой сорт? Почему?
 —")
• Евге ника (от др. греч. εὐγενής — «хорошего рода, благородный» ) — учение о селекции применительно к человеку, а также о путях улучшения его наследственных свойств. Учение было призвано бороться с явлениями вырождения в человеческом генофонде. • Отцом же евгеники, поставившей ее на научную основу, стал Френсис Гальтон (двоюродный брат Чарльза Дарвина) в 1869 году. Проанализировав родословные сотни талантливых людей, он пришел к выводу: гениальные способности наследуются. По его мнению, подтверждала право англосаксонской расы на мировое господство • В древней Спарте селекцию людей проводили более кардинально, уничтожая младенцев, не обладающих физическими качествами, необходимыми для будущего воина.

• На Международном Конгрессе по вопросам евгеники, который проходил в Нью Йорке в 1932 году, один из учёных специалистов евгеников прямо заявил следующее: • «Нет никакого сомнения, что если бы в Соединённых Штатах закон о стерилизации применялся бы в большей мере, то в результате меньше чем через сто лет мы ликвидировали бы по меньшей мере 90 % преступлений, безумия, слабоумия, идиотизма и половых извращений, не говоря уже о многих других формах дефективности и дегенерации. Таким образом, в течение столетия наши сумасшедшие дома, тюрьмы и психиатрические клиники были бы почти очищены от своих жертв человеческого горя и страдания. »

• Одни из самых жестоких евгенических законов существовали в Северной Каролине. К примеру, стерилизация делалась автоматически всем людям, чей IQ был ниже 70. Также поощрялась стерилизация среди бедняков — за эту операцию им даже платилась премия в 200 долларов. • В Европе такая кастрация была впервые осуществлёна в 1925 году в Дании, по решению суда. • С 1934 по 1976 годы программа принудительной стерилизации «неполноценных» осуществлялась в Швеции. Схожие законы действовали в Норвегии и Финляндии, а также Эстонии и Швейцарии. • В нацистской Германии (1933— 1945) принудительной стерилизации подлежали все «неполноценные лица» : евреи, цыгане, люди с отклонениями в развитии, душевно больные, коммунисты и т. д. Затем было принято решение о большей сообразности их физического уничтожения


• В начале ХХ века Первый мир был обуреваем страстью к переделке человека. Результаты не замедлили появиться: русско французский субботник (русский субэтнос, исповедовавший иудаизм) Воронов и австриец Штейнах с целью омоложения провели сотни пересадок яичек обезьян людям. Но в 1940 е по этическим соображениям эти опыты были запрещены. • Сергей Воронов за операцией • «Собачье сердце» Михаила Булгакова

• Евгеника • «За» и «Против»

• Эту девушку зовут Мелани Гайдос, и она ворвалась в мир моды стремительно, эпатируя, воодушевляя и разрушая глупые стереотипы. Она смогла добиться популярности, и сделала это, несмотря на свой генетический недуг. Читайте подробнее на FB. ru: http: //fb. ru/post/culture/2017/7/11/13859





Д/З п. 27 Дополнительно сообщения по теме
Мутагены — факторы окружающей среды, которые способны вызывать ошибки при нормальной репликации ДНК, что ведет к спонтанным мутациям. Ошибки в репликации ДНК зависят от температурных условий, pH, состава среды. Особенно мощными мутагенными факторами являются ультрафиолетовые и ионизирующие излучения. К мутагенам химической природы относятся аналоги азотистых оснований, отдельные красители акридинового ряда, алкилирующие соединения, некоторж антибиотики, гидроксиламин, уретан и азотистая кислота. [c.61]
Химическое изменение оснований. Некоторые мутагенные вещества действуют путем химического изменения содержащихся в ДНК оснований, что приводит к ошибкам репликации. Вполне понятное изменение вызывает нитрит. Азотистая кислота дезаминирует аденин, гуанин или цитозин без разрыва или каких-либо других изменений полинуклеотидной цепи. В результате замещения аминогруппы гидроксильной группой аденин превращается в гипоксантин и спаривается с цитозином вместо тимина, что приводит к мутации АТ СС. Если цитозин дезаминируется в урацил, то он спаривается с аденином вместо гуанина, и это ведет к мутации СС -АТ. Будучи превращен в ксантин, гуанин по-прежнему спаривается с цитозином, т. е. дезаминирование С не вызывает мутации. Гидроксиламин вступает в реакцию главным образом с цитозином и изменяет его так, что тот спаривается с аденином значит, он тоже вызывает мутации СС ТА. [c.444]
Ошибки репликации ДНК приводят к мутациям [12] [c.128]
Мутации как ошибки репликации ДНК. Данные, полученные на человеке, свидетельствуют о существовании тесной связи между мутациями и клеточными делениями. Импульсом к изучению этой проблемы послужила гипотеза о механизме возникновения точковых мутаций Уотсона и Крика (рис. 5.26) [1347]. Важную роль в этом сыграли и ранние исследования на микроорганизмах, из которых следовало, что многие спонтанные мутации действительно возникают во время репликации ДНК в результате ошибочной вставки неправильного нуклеотида, приводящей к появлению в будущих клеточных поколениях новой, отличающейся пары оснований. В последние десятилетия получено до удивления мало новых данных о механизмах возникновения спонтанных мутаций, что очень сильно контрастирует с громадным объемом имеющихся в настоящее [c.191]
Заметим, что различие по длине рестрикционных фрагментов между родителями и детьми может возникать не только в результате точковых мутаций в сайте узнавания, но и в результате ошибки репликации или кроссинговера. Ожидается, что эти события происходят чаще, чем точковые мутации. Среди 27 индивидов, у которых проанализировано 240 полос, выявлена такая полоса, которой не было ни у одного из родителей, отсюда скорость мутирования составляет 1/240, что по крайней мере на 4 порядка выше, чем скорость мутирования для точковых мутаций (разд. 2.3.3.9). [c.290]
Передача наследственной информации в неискаженном виде — важнейшее условие выживания как каждого конкретного организма, так и вида в целом. Следовательно, в ходе эволюции должна была сформироваться система, позволяющая клетке исправлять нарушения ДНК, вызванные ошибками репликации или повреждающими воздействиями окружающей среды. Подсчитано, что в результате повреждений, обусловленных этими причинами, в геноме клеток зародышевой линии человека происходит в среднем шесть нуклеотидных замен в год. По-видимому, в соматических клетках за год происходит примерно такое же число мутаций. [c.79]
В работе [59] было показано, что 95% мутаций, индуцируемых АП в Е.соЫ, относятся к транзициям А—Т — -> Г — Ц, т. е. преимущественно возникают ошибки репликации. [c.34]
Данные таблицы указывают, что в Si состоянии А и Т (У) легче, а Ц и Г труднее переходят в редкие формы, чем в состоянии Sq. Видно также, что в случае амино-иминной таутомерии А более склонен к таутомеризации, чем Ц лактам-лактимные превращения Т (У) более вероятны, чем Г. Таким образом, в состоянии Si положение противоположно тому, что имеется в So- Поэтому при действии УФ-света ошибки репликации чаще всего приводят к транзициям А—Т Г—Ц (рис. 3, а и б), а ошибки включения — к транзициям Г —Ц- А—Т (рис. 3, ж и з). Рассмотренный механизм объясняет возникновение мутаций, отнесенных к транзициям А — Т — Г — Ц [177]. [c.54]
ДНК-полимеразы проверяют комплементарность каждого нуклеотида матрице дважды один раз перед включением его в состав растущей цепи и второй раз перед тем, как включить следующий нуклеотид. Очередная фосфодиэфирная связь образуется лишь в том случае, если последний (З -концевой) нуклеотид затравки комплементарен матрице. Если же на предыдущей стадии полимеризации произошла ошибка (например, из-за того, что нуклеотид в момент полимеризации находился в необычной таутомерной форме), то репликация останавливается до тех пор, пока неправильный нуклеотид не будет удален. Некоторые ДНК-полимеразы обладают не только полимеризующей, но и 3 -экзонуклеазной активностью, «Которая отщепляет не спаренный с матрицей нуклеотид затравки. После чего полимеризация восстанавливается, от механизм, коррекция, заметно увеличивает точность работы ДНК-полимераз. Мутации, нарушающие З -экзонуклеазную активность ДНК-полимеразы, существенно повышают частоту возникновения прочих мутаций. Напротив, мутации, приводящие к усилению экзонуклеазной актив- ности относительно полимеризующей, снижают темп мутирования Генетического материала. [c.47]
Спонтанные генные мутации определяются ошибками при репликации ДНК, возникающими вследствие теплового движе-иия атомов и молекул. Очевидно, что ошибки транскрипции и трансляции не наследуются. [c.283]
Если ошибка синтеза не устраняется системами репарации, то неизбежна деформация дуплекса и искажение генетической программы. Такие сохраняющиеся при репликации изменения ДНК носят название мутации. Они могут быть спонтанными и индуцированными. Частота спонтанных мутаций невелика и составляет всего 10 —10 на клетку. В основном имеют место мутации, обусловленные действием внешних факторов физических (радиация), биологических (вирусы) и чужеродных химических веществ на генетический аппарат клеток. Наиболее многочисленными и опасными являются мутагены окружающей среды. Загрязнение воды и воздуха различными химическими отходами промышленных предприятий, химическими средствами защиты растений отрицательно сказывается на генетической программе всех живых организмов. В последние годы установлено, что ряд пищевых красителей, стабилизаторов и вкусовых добавок обладает выраженной мутагенной активностью, что привело к значительному ужесточению требований, связанных с применением химических веществ в пищевой промышленности. Многие лекарственные вещества также воздействуют на генетический аппарат клеток и должны подвергаться специальным генетическим испытаниям. [c.455]
Из уровня спонтанных мутаций у бактерий в расчете на одно поколение рассчитано, что вероятность одной репликационной ошибки при синтезе ДНК составляет порядка 10 . Эту величину можно рассматривать как отношение скоростей реакций правильной репликации [c.194]
Как обсуждалось в гл. 13, наследственная информация, заключенная в нуклеотидной последовательности ДНК, сохраняется неизменной благодаря действию сложных метаболических механизмов, обеспечивающих осуществление репликации и репарации. Мутации могут быть результатом ошибки на любом из многочисленных последовательных этапов этих процессов. Мутагенные факторы способны изменять как непосредственно структуру ДНК, так и структуру ферментов, прямо или косвенно участвующих в соответствующих метаболических процессах. Для понимания механизмов мутаций требуется знание нуклеотидной последовательности гена дикого типа и мутантного гена. Без этого невозможно понять связь между изменениями, происходящими в структуре ДНК и действием конкретных факторов или условий среды, вызывающих мутации. Современные методы клонирования генов сделали возможным прямое определение нуклеотидной последовательности ДНК. Однако еще совсем недавно при изучении молекулярной природы мутаций приходилось анализировать аминокислотные замены в белках, синтезируемых мутантными генами, а затем с помощью таблиц генетического кода выявлять изменения в нуклеотидной последовательности. [c.8]
Связь между ДНК, менделевскими факторами и признаками организма не столь проста, как представлялось прежде, но под наблюдения классических менделистов можно тем не менее подвести химическую основу. Установленные ранее явления доминантности и рецессивности рассматриваются теперь в таких аспектах, что можно без труда показать связь этих явлений с определенными химическими реакциями (разд. 2.1.2). Эпистаз (разд. 2.1.2) можно интерпретировать в понятиях взаимодействующих генов и генных продуктов. Нетрудно, например, представить себе, как может происходить химическое воздействие на экспрессию на уровне как самого гена, так и любого из ряда последовательных генных продуктов химическими факторами, содержащимися во внутренней и во внешней среде. Наконец, мутацию можно интерпретировать как ошибку в репликации ДНК. Такая ошибка может ограничиваться замещением одного нуклеотида другим (точковая мутация) или же выражаться в структурной перестройке целых хромосом (хромосомные мутации, в том числе нехватки, удвоения, транслокации и инверсии). [c.38]
Если УФ-свет вызывает мутацию гена, ответственного за биосинтез ДНК-полимеразы или других ферментов репарирующей системы, то возникшие мутантные клетки характеризуются высокой частотой спонтанных мутаций, которые не связаны с действием внешних или внутренних мутагенных факторов, а обусловлены ошибками в ходе репликации. В связи с этим в генетике возникло представление о гене, контролирующем частоту естественных мутаций,— гене-мута-торе. [c.311]
Устойчивость процесса отбора по отношению к систематическим помехам зависит прежде всего от уровня помех, т. е. от вероятности мутаций в единицу времени и вероятности ошибочной репликации. Следует учитывать также различие в характере повреждений, вызываемых помехами. При ошибочной редупликации (или в результате мутаций) могут возникнуть как летальные варианты, т. е. гиперциклы с весьма малой репродуктивной способностью, так и гиперциклы, способные к дальнейшему развитию, но отличающиеся от исходного. Характер повреждений зависит от стадии развития. На первой стадии (до возникновения адаптеров) все мутации и ошибки ведут к равноправным вариантам, летальных вариантов нет. На второй стадии, после образования адаптеров, но до возникновения единого кода летальные мутации существенной роли не играют, поскольку сводятся к уменьшению скорости репродукции. [c.36]
На наших глазах очередной кризис возник и в молекулярной генетике. Совсем недавно доминировали представления о необычайной точности и стабильности механизмов репликации и процессинга РНК, в результате чего мутации (ошибки репликации) возникают крайне редко, с частотой порядка 10 10 за одно поколение. Немало приводилось убедительных доводов в пользу того, что только такая надежность репликации ДНК и обеспечивает саму возможность прогрессивной эволюции — при учащении мутационного процесса отбор-де не справился бы с задачей поддержицання упорядоченности организации и совершевствовавяя [c.4]
По происхождению мутации делятся на спонтанные (неконтролируемые) и индуцированные (контролируемые). Первые возникают в результате неконтролируемого влияния каких-то естественных факторов (радиация, температура и т. д.). Направленное использование мутагенов приводит к возникновению индуцированных мутаций. Многими экспериментами четко показано, что мутации возникают независимо от условий среды обитания, т. е. не направленно. Мутации возникают в основном как ошибки репликации ДНК. Выделяют следующие типы мутаций перестройка хромосом, перестройка генома клетки грибов и водорослей (полиплоидия, гаплоидия, гетероплоидия), внутригенные изменения (прямые мутации, реверсии, обратные мутации). [c.102]
В качестве альтернативного подхода к измерению скоростей мутации А. Portner и соавт. [198] использовали моноклональные антитела и сравнили скорости мутации в отношении к резистентности антител к вирусам гриппа, везикулярного стоматита и Синдбис. Для всех трех вирусов скорости мутации составили около на репликацию, тогда как скорость ошибки репликации ДНК Е. соИ оценивалась как 10 —10 ° [63]. [c.190]
Спонтанные мутации возникают случайно, т.е. в любой. момент любой ген может претерпеть изменения. Причинами спонтанного мутационного процесса являются многочисленные факторы экзогенной и эндогенной природы, в том числе постоянное воздействие па организм человека мутагенов химической, биологической и физической природы (например, естественный фон облучения, действие вирусов) ошибки репликации ДНК, которые копируются и накапливаются в ряду клеточных поколений нарушение функционирования репаративных систем действие экзогенных метаболитов физи0 ю1ическ0е состояние и возраст организма. Спонтанные мутации могут возникать как в половых, так и в соматических клетках на генном, хромосомном и геномном уровнях. [c.125]
Разные аллели одного и того же Г. возникают благодаря мутациям-илслецуемьш изменениям в структуре исходного Г. В норме Г. чрезвычайно стабилен и при удвоении хромосом во время репликации ДНК воспроизводится совершенно точно вероятность ошибки не превышает 10″ . Мутации происходят редко и обычно влекут за собой неблагоприятные последствия для организма, т. к. нарушается его способность синтезировать нормальный белок. Однако в целом это явление играет положит, роль накопление редких полезных мутаций создает основу генетич. изменчивости, необходимой для эволюции. [c.517]
Равновесие между созидательными возможностями выбора среди специфических оснований в ДНК (созидательные мутации) и точностью синтеза белков (поддерживающих жизнь организма) является основой эволюции. Ферменты, которые заряжают тРНК специфической аминокислотой, обладают очень низкой вероятностью ошибки, порядка 1 Ю» для гомологичных аминокислот. При репликации точность даже выше, и величина ошибки редко превышает 1 на 10. [c.212]
Приближенная модель репликации ДНКизображена на рис. 2.11. Из приведенной схемы видно, что репликация точно воспроизводит прежнюю (исходную) структуру ДНК. Но если произошла ошибка в процессе копирования (мутация), то она будет с предельной точностью копироваться при последующих репликациях изменившейся ДНК. Показано, что участки ДНК, содержащие скопления нуклеотидов, обладают повышенной склонностью к спонтанным мутациям [22]. [c.94]
Для нормального функционирования аппарата исправления ошибок, связанных с включением неправильных нуклеотидов, необходимо располагать механизмом, позволяющим отличать новосинтезированную цепь ДНК от родительской матричной цепи. В противном случае с вероятностью 1/2 будет происходить исправление нуклеотида в родительской цепи, приводящее к закреплению потенциально мутагенной ошибки, допущенной ДНК-полимеразой. Вероятно, для установления различий между родительской и дочерней цепями ДНК в Е. соИ используется метилирование аденина в последовательности GAT . Эта палиндром-ная последовательность обычно метилирована в обеих цепях родительской ДНК. При полуконсервативной репликации метилированной ДНК образуется дочерняя ДНК, в которой одна цепь, пришедшая от родительской ДНК, метилирована, а новообразованная цепь в течение некоторого времени после выхода из области репликативной вилки остается неметилированной. Следует заметить, что метилирование новообразованной цепи ДНК осуществляется ферментом, отличным от метилаз, входящих в систему рестрикции—модификации, обсуждавшуюся в гл. 9. Бактерии dam , дефектные по метилированию аденина в результате нарушения синтеза соответствующей метилазы характеризуются повышенной частотой спонтанных мутаций, что подтверждает гипотезу об участии метилазы dam в системе исправления ошибок репликации. [c.123]
Анализ приведенных выше результатов дает возможность написать для преобладающих таутомерных форм оснований нуклеиновых кислот формулы, изображенные на фиг. 55. Минорные таутомерные формы, возможно, играют существенную роль в возникновении спонтанных мутаций, поскольку спаривание несоответствующих оснований (см. гл. ХУП1) должно привести к ошибке при включении оснований и при последующей репликации цепи. Можно показать, что если скорость включения основания в цепь нуклеиновой кислоты меньше скорости перехода минорного таутомера в доминирующую форму, то скорость спонтанных мутаций, обусловленных данным основанием, приблизительно равна константе равновесия между минорным и доминирующим таутомерами. К сожалению, для азо- [c.308]
Задание 189. Напишите программу для моделирования самоорганизации ДНК в качестве примера самоорганизуюшихся систем. Используйте для этого следующую простую модель. Пусть имеется 100 молекул ДНК, состоящих из 12 нуклеотидов четырех видов (их обозначим буквами А, Т, С и G). Последовательность нуклеотидов в этих 100 молекулах ДНК случайная. Назовем одну из последовательностей идеальной она имеет некоторые преимущества перед остальными. Из 100 молекул ДНК в результате репликации получается еще 100 молекул. Однако при репликации встречаются ошибки (мутации), например в количестве 1%. Теперь из 200 молекул 100 погибает. При этом имеет значение преимущество, которым обладают молекулы с последовательностью нуклеотидов, похожей на идеальную . (Например, при каждом совпадении нуклеотида и его положения в цепи с идеальной последовательностью вероятность гибели уменьшается в два раза.) Процессы репликации и гибели протекают очень быстро. В конце концов все молекулы ДНК должны получить идеальную последовательность нуклеотидов, хотя вероятность ее образования в результате случайного процесса составляет 1 16777216. Что будет, если мутации будут возникать чаще или реже [c.330]
Теперь мы уже вполне подготовлены к тому, чтобы приступить к вопросу, поставленному в гл. VU, а именно к вопросу о молекулярном механизме возникновения тех изменений в последовательности нуклеотидов ДНК, которые приводят к мутациям. Действительно, исследование характера возникновения мутаций Т-четных фагов с использованием методов генетического анализа с высоким разрешением дает большие возможности для проникновения в природу мутационного процесса. Использование фагов имеет еще одно важное преимущество по сравнению с ис-лользованием бактерий. Мутации фаговой ДНК можно изучать как в том случае, когда она находится в состоянии покоя вне клетки в составе инфекционной фаговой частицы, так и когда она находится в реплицирующемся, внутриклеточном, вегетативном состоянии. Уже самые первые исследования Херши и Лурия показали, что частота спонтанных мутаций в покоящейся ДНК очень мала — столь мала, что в течение многих лет считалось (как потом оказалось, ошибочно), что внеклеточные фаговые частицы вообще не мутируют месяцами и даже годами. Таким образом, новые мутации появляются в основном во время вегетативного размножения фага в клетке-хозяине. Рассмотрим следующий пример. Культуру Е. oli заражают препаратом фага Т2/- с титром 10 частица/мл. Фагу дают размножиться в течение нескольких циклов, пока все бактерии в культуре не подвергнутся лизису, а титр фага не достигнет величины 10 частица/мл. Оказывается при этом, что с каждым циклом размножения доля г-мутантов во всей популяции фагов увеличивается (примерно с 10″ в начале до 10 в конце). Следовательно, мутанты фага возникают в результате ошибок копирования при внутриклеточной репликации его генетического материала. Репликация ДНК родительского фага является очень точным процессом. И все же при репликации иногда происходит ошибка, порождающая в одной из вегетативных реплик изменение последовательности нуклеотидов, или мутацию. Мутантная реплика генетического материала включается затем при созревании в инфекционную фаговую частицу, которая в свою очередь заражает новую бактериальную клетку. В этой клетке очень точно копируется уже измененная информация, содержащаяся в мутантной частице поэтому все потомство такой частицы оказывается тоже мутантным. Поскольку репликация ДНК вегетативного фага происходит в соответствии с постулированным Уотсоном и Криком полуконсервативным механизмом, размножение фагового генома можно рассматривать как процесс бинарного деления и с точки зрения статистического анализа совершенно аналогичным процессу размножения генома бактерий. Следовательно, уравнение, связывающее долю мутантных особей п среди общего числа N потомков одного исходного родителя, возникших после g генераций, с частотой мутаций а [c.315]
Если элементарное мутационное событие представляет собой [включение неправильного нуклеотида в определенный участок синтезируе-мой полинуклеотидной реплики и если ДНК вегетативного фага реплицируется в соответствии с полуконсервативным механизмом Уотсона и Крика, то мы можем предсказать такую особенность вновь рождаюш егося мутантного генома, которую без знания молекулярной основы процесса мутирования вообще невозможно было бы предвидеть. Предположим, что во время синтеза цепи-реплики происходит одна из редких ошибок копирования, например остаток тимина в родительской цепи незаконно спаривается с гуанином, а не с аденином. В результате этого мутагенного акта репликации возникает двойная спираль с исходной ин-формацией в старой (родительской) цепи и мутантной информацией в цепи, синтезированной заново (фиг. 160). При следующем цикле репликации комплементарные нити этой мутантной молекулы вновь разъединяются и каждая из них, функционируя как матрица, синтезирует новую комплементарную цепь. В результате появляется одна двойная спираль ДНК, несущая мутантную информацию в обеих цепях, и одна немутантная двойная спираль. Исходная мутантная молекула ДНК представляет собой, следовательно, гетеродуплексную гетерозиготу, которая несет в одном участке два аллеля — мутантный и немутантный, по которым при следующем цикле репликации происходит расщепление. Можно ожидать, что во время внутриклеточного размножения фага некоторые молекулы ДНК фага с мутацией, возникшей в результате ошибки копирования при последней репликации, будут извлечены из вегетативного фонда фага и войдут в состав зрелых инфекционных частиц. Эти частицы и будут мутационными гетерозиготами. [c.325]
Во-первых, исходным повреждением, отвечающим за потенциальный мутагенный эффект, должны быть тиминовые димеры. Во-вторых, процессом, превращающим потенциальные изменения в окончательные мутации, не может быть репарация за счет иссечения и заполнения (она могла бы приводить к мутации, если бы, например, точность репарационной репликации, изображенной на фиг. 187, была невелика и допускала ошибки копирования). Можно заключить, следовательно, что мутация вызывается нерепарированными тиминовыми димерами в тех клетках, которые выжили, несмотря на наличие такого неисправленного п овреж- [c.382]
Другой путь возникновения транзиций-это случаи ошибочного спаривания, приводящие к возникновению неканонических пар и, следовательно, к дефектам в уотсон-криковской спирали. В нормальном цикле репликации такая ошибка может случайно произойти вследствие включения неправильного основания. Спонтанная частота ошибок определяется прежде всего точностью фермента ДНК-полимеразы, отвечающей за репликацию (см. гл. 32). Существует также более ограниченный репара-тивный синтез ДНК, который активируется в результате генетической рекомбинации или повреждения ДНК (см. гл. 34). Различные системы репарации характеризуются разной частотой ошибок. Например, одна из репара-тивных систем Е. соИ особенно часто делает ошибки, и, следовательно, ее активация может стимулировать образование мутаций. Мы не располагаем достаточной информацией о частоте возникновения мутаций такого рода. [c.38]
Что представляет собой механизм появления ошибок Можно предположить, что определенный компонент пути репарации обусловливает продолжение репликации за сайтом повреждения. Когда ДНК-полимераза минует тиминовый димер, она включает неправильные основания и это приводит к появлению мутации. Существуют доказательства того, что для индукции ошибок необходимо присутствие ДНК-полимеразы III, обычной репликазы. Следовательно, рассматриваемая функция действует согласованно с нормальным реплика-ционным аппаратом. Мутации в гене, получившем название итиС, устраняют УФ-индуцируемый мутагенез, но не нарушают какие-либо известные ферментативные функции. Вероятно, продукт этого гена, итиС, служит компонентом системы, продуцирующей ошибки. [c.440]
Модель Уотсона-Крика позволяет представить себе, как может удваиваться нативная молекула ДНК, образуя две одинаковые дочерние молекулы. Поскольку две цепи ДНК комплементарны, каждая из них при расплетании двойной спирали может служить матрицей для синтеза новой комплементарной цепи. Последовательность оснований во вновь синтезируемой цепи будет определяться спецификой водородных связей между основаниями цепи-щаблона и вновь образуемой цепи (рис. 4.13). Таким образом, генетическая информация, содержавшаяся в последовательности пар оснований родительской молекулы, будет полностью воспроизведена в двух дочерних молекулах. Более того, если в процессе удвоения ДНК произошла ошибка и какой-то нуклеотид во вновь образуемой цепи выпал или оказался некомплементарным исходному, то это может изменить информационное содержание молекулы, причем можно ожидать, что эта ошибка будет передана дочерним молекулам ДНК в следующих поколениях. Такая замена пары нуклеотидов может обладать свойствами генетических мутаций. Таким образом, модель структуры ДНК Уотсона и Крика объясняет как способность генов к самоудвоению (репликации), так и их информационные свойства. [c.107]
Несмотря на корректорские функции, присущие ДНК-полимеразам Е. соИ, некоторые нуклеотиды оказываются все же ошибочно включенными в новообразованную цепь ДНК. Их присутствие делает возможным возникновение спонтанных мутаций, в том случае если ошибки не будут исправлены до начала следующего цикла репликации. Свидетельства в пользу существования пострепликационных систем исправления ошибок, или репарации, были получены при изучении таких явлений, как [c.122]
Спонтанные трацзиции могут происходить при репликации ДНК вследствие таутомеризации, т. е. изменения положения протона, меняющего химические свойства молекулы. Таутомеризация в нуклеотидных основаниях меняет их способность образовывать водородные связи, так что аденин приобретает свойства гуанина, гуанин-аденина, цитозин-тимина, а тимин-цитозина (рис. 20.2). Мутагенная активность 5-бром-урацила, аналога тимина, в котором метиловая группа замещена атомом брома, обусловлена таутомеризацией, связанной с большим, нежели у метиловой группы, сродством к электрону атома брома по сравнению с метиловой группой (рис. 20.3). Индуцируемые 5-бромура-цилом мутации могут обусловливаться либо ошибками при включении, [c.9]
Обновлено: 06.06.2023
Генные мутации присутствуют в каждом человеке. Ученые определили среднее число мутантных генов и скорость возникновения мутаций, которая составляет 100–200 мутаций на поколение.
При слове «мутация» в сознании возникают либо страшноватые образы двухголовых козочек, либо фантастические сверхсущества из фильма «Люди Х». Однако на самом деле в мутациях нет ничего необычного. Без преувеличения можно сказать, что мы все мутанты. Вопрос только в том, какой процент мутировавших генов содержит наша ДНК.
(англ. John Burdon Sanderson Haldane; сокращённо Дж. Б. С. Холдейн, англ. J. B. S. Haldane; 5 ноября 1892, Оксфорд,Оксфордшир, Великобритания — 1 декабря 1964, Бхубанешвар, штат Орисса, Индия) — английский биолог (генетик.
Первая попытка рассчитать темпы мутации генома человека была предпринята в 1935 году одним из отцов современной генетики англичанином Джоном Холдейном. При обследовании мужчины, больного гемофилией, он пришел к выводу, что только в одном из 50000 случаев мутация гена вызывает гемофилию. Это соответствует мутации одного из 25 млн нуклеотидов генома. После Холдейна скорость возникновения мутаций пытались определить, сравнивая ДНК человека и шимпанзе, однако, конечно, точные данные не были получены.
Однако возможности современной генетики позволяют получить точные данные о скорости мутаций — их приводит интернациональная группа из 16 ученых в работе, опубликованной в Current Biology. Они показали, что приблизительные данные, полученные Холдейном 70 лет назад, оказались не так уж далеки от реальности.
стойкое (то есть такое, которое может быть унаследовано потомками данной клетки или организма) изменение генотипа, происходящие под влиянием внешней или внутренней среды. Процесс возникновения мутаций получил название мутагенеза.
Каждый человек является носителем одной мутации в каждых 15–30 млн нуклеотидов.
Для расчета скорости возникновения мутаций авторы работы изучили фрагмент ДНК двух мужчин из китайской деревни, чьи предки несколько сот лет жили в этом же регионе. Общий предок этих мужчин отделен от них 13 поколениями и жил около 200 лет назад. Для чистоты эксперимента ученые исследовали фрагмент мужской Y-хромосомы. Она состоит из 10149085 пар нуклеотидов и передается от отца к сыну в неизменном виде (Y-хромосома у женщин отсутствует). Используя современные методы расшифровки генома, ученые установили, что 10149073 нуклеотидные пары у мужчин не отличимы, то есть всего было локализовано 12 мутаций. Восемь из них при дальнейшем исследовании оказались возникшими уже в клетках взрослого человека в результате их жизнедеятельности, а четыре оказались истинными мутациями, возникшими из-за «ошибки» при передаче генетического материала от отца к сыну.

Приняв эти данные за средние по всему геному и пересчитав их на общее количество генов (полный геном содержит более трех миллиардов нуклеотидов) и 13 поколений, разделяющих мужчин,
ученые рассчитали темпы появления мутаций в человеческом геноме: 100–200 мутаций на поколение.
Большая часть этих мутаций являются безвредным и, в принципе, не заметными для человека, для его организма и здоровья. Однако в редких случаях мутации могут приводит либо к врожденным тяжелым болезням — например, к раку или диабету, либо вносить «усовершенствования» в организм, делая его более стойким.

Интерес к возникновению мутаций и темпам их роста отнюдь не является праздным. Главная их роль — отнюдь не возникновение неизлечимой болезни у какого-либо конкретного человека. Мутации являются необходимым для движения эволюции материалом. Именно они дают генетическое разнообразие, которое позволяет живому миру двигаться вперед. Конечно, на одном-двух поколениях проследить эволюцию невозможно, однако именно мутации дают изменение генома, которое, в случае выгодности его для организма, повышает его стойкость. Если мутация выгодна, то именно носители такого мутантного гена выживают поколение за поколением, в конце концов скрещиваются, и мутация закрепляется уже как системное изменение.
Поэтому изучение скорости и механизма возникновения мутаций может позволить распутать цепь эволюции с конца, как клубок, и прояснить «белые пятна» в истории происхождения видов.
X Международная студенческая научная конференция Студенческий научный форум — 2018

ГЕНОМНЫЕ ХРОМОСОМНЫЕ МУТАЦИИ В ГАМЕТОГЕНЕЗЕ И ЭМБРИОГЕНЕЗЕ ЧЕЛОВЕКА
Текст работы размещён без изображений и формул.
Полная версия работы доступна во вкладке «Файлы работы» в формате PDF
Цель: понять причины возникновений хромосомных мутаций в гаметогенезе и эмбриогенезе человека.
1. Понять, что такое хромосомные мутации.
2. Разобрать причины их возникновений.
3. Выяснить к каким патологиям приводят хромосомные мутации.
1.Что такое геномные хромосомные мутации. 4
2. Геномные мутации. 5
2.1 Полплоидия. 6
2.2 Анэуплоидия. 7
Хромосомные мутации. 8
Список использованной литературы. 14
В современной медицине при изучении болезней человека важное значение имеют наследственные заболевания, связанные с нарушениями целостности генетической информации в геноме человека, вызванными различными факторами.
Геном — наследственный аппарат клетки, содержащий весь объём информации, необходимый для её существования в условиях среды и передачи наследственных признаков последующему поколению.
Геномика – наука, изучающая геном человека и геномы в целом.
Медицинская генетика – одно из направлений геномики человека, система знаний о роли генетических факторов в патологии человека и система методов диагностики, лечения и профилактики наследственной патологии в широком смысле.
Эта наука играет важную роль в профилактической медицине, позволяя посредством различных методов генной терапии предупредить рождение больного ребёнка в семье с наследственной патологией.
Генная терапия предполагает лечение самых разнообразных, а не только наследственных болезней с помощью введения больному генов, играющих ключевую роль в патогенезе соответствующих заболеваний.
Медицинская генетика имеет важное значение в профилактике и лечении любых заболеваний, связанных с какими либо генетическими нарушениями, не только наследственного характера.
1. Что такое геномные хромосомные мутации?
Материальной субстанцией наследственности являются молекулы ДНК и, в частности гены – транскрибируемые фрагменты ДНК, кодирующие белки и разнообразные молекулы РНК (рРНК, тРНК, регуляторные и другие РНК). Изменчивость определяется существованием различных состояний генов или аллелей. При этом нормальная изменчивость связана с присутствием у разных индивидуумов нормальных вариантов гена, а патологическая изменчивость – с наличием множества мутантных аллелей или мутаций. Носители хромосомных аномалий, доминантных мутаций или гомозиготы по рецессивным мутациям называются мутантными особями или мутантами. Мутации называются «легкими» или «тяжелыми», если они ассоциированы с мягким или тяжелым течением заболевания соответственно.
Мутации бывают геномными, хромосомными или генными. В общем случае, геномные и хромосомные мутации приводят к тяжелым патологическим состояниям, часто несовместимым с жизнью.
К геномным мутациям относятся увеличения полного набора хромосом – полиплоидии, или изменения количества хромосом одной пары – анеуплоидии. У человека описано два вида полиплоидий – триплоидии и тетраплоидии – трех- и четырехкратное увеличение числа гаплоидного набора. Подобные аномалии встречаются только у спонтанных абортусов или мертворожденных. [4 c.55]
Полиплоидия (эуплоидия) – геномная мутация, вызванная добавлением целого гаплоидного набора хромосом как в результате ошибок в процессе мейоза, так и при нарушении митоза.
Гаметы и соматические клетки с увеличенным набором хромосом, кратным их гаплоидному числу, называют полиплоидными. Приставки три-, тетра- и т. д. указывают, во сколько раз увеличено число хромосом, т. е. степень плоидности:3n-триплоид, 4n-тетраплоид, 5n-пентаплоид и т. д.
У растений полиплоидия встречается гораздо чаще, чем у животных. Относительная редкость полиплоидии у животных объясняется тем, что увеличение числа хромосом значительно повышает вероятность ошибок при образовании гамет в мейозе.
У человека описаны триплоидные и тетраплоидные организмы. Частота их возникновения низка. Они обнаруживаются среди спонтанно абортированных эмбрионов или плодов и у мертворожденных. Продолжительность жизни новорожденных с такими нарушениями — несколько дней.
Триплоидия может быть обусловлена нарушением мейотического расхождения всего набора хромосом в мейозе женских (отсутствие первого мейотического деления ооцита) или мужских половых клеток. В результате либо яйцеклетка, либо сперматозоид оказываются диплоидными. В качестве механизма триплоидии рассматривают также возможность оплодотворения яйцеклетки двумя сперматозоидами. В том случае, когда триплоидия обусловлена отцовским диплоидным набором хромосом, возникает пузырное перерождение плаценты, так называемый пузырный занос, препятствующий стабильному поступлению питательных веществ от матери к ребенку, а следовательно, его нормальному развитию.
Гораздо больший интерес у учёных-генетиков вызывают заболевания, причиной которых является анэуплоидия.
Анэуплоидия может выражаться в появлении в дочерних клетках добавочной хромосомы (n+1),(2n+1) или в нехватке какой-либо хромосомы (n-1),(2n-1) и т.д. Анэуплоидия может возникнуть, если в анафазе I мейоза гомологичные хромосомы одной или нескольких пар не разойдутся. В этом случае оба члена пары направляются к одному и тому же полюсу клетки, и тогда разделение гомологичных хромосом в анафазе II может привести к образованию гамет, содержащих на одну или несколько хромосом больше или меньше, чем в норме. Это явление известно под названием нерасхождения. Когда гамета с недостающей или лишней хромосомой сливается с нормальной гаплоидной гаметой, образуется зигота с нечетным числом хромосом: вместо каких-либо двух гомологов в такой зиготе их может быть три или только один.
Зигота, в которой число хромосом меньше диплоидного, обычно не развивается, но зиготы с лишними хромосомами иногда способны к развитию. Если это происходит у животных, то из таких зигот в большинстве случаев развиваются особи с резко выраженными аномалиями. У человека наиболее ярким примером нерасхождения хромосом является синдром Дауна, трисомия по 21-й паре хромосом.
Возможно также нерасхождение мужских и женских половых хромосом, которое приводит к анэуплоидии, влияющей на вторичные половые признаки и фертильность, а иногда и на умственные способности.
Хромосомные мутации, в свою очередь, могут быть числовыми (анеуплоидии) или структурными, то есть затрагивать число хромосом или их структуру. Наиболее частыми числовыми аномалиями являются моносомии – отсутствие одной из гомологичных хромосом и трисомии – существование добавочной третьей копии одной из гомологичных хромосом, причем эта добавочная хромосома может быть как материнского, так и отцовского происхождения. Трисомии найдены не для всех хромосом, и наиболее частыми из них являются синдромы Дауна, Эдвардса и Патау – трисомии по 21, 18 и 13 хромосомам соответственно. Иногда количество добавочных хромосом может быть еще больше, эти аномалии называются полисомиями. Моносомии и полисомии описаны, главным образом, для половых хромосом. Другие геномные мутации несовместимы с жизнью и приводят к ранней эмбриональной гибели.[4 c.56]
Синдром Шерешевского Тернера-заболевание ,вызванное моносомией по Х-хромосоме(45 хромосом = 44 аутосомы + ХО).
В период созревания гамет наблюдаются случаи нерасхождения половых хромосом (в I, II или обоих делениях созревания).
Гаметы несут не 22 аутосомы + 1половую хромосому (Х или У), а возникает нарушение парности хромосом. Моносомия Х зависит исключительно от отца.
Р ♀ 44 аутосомы + ХХ → ♂ 44 аутосомы + XY
Гаметы 22 аутосомы + Х 22 аутосомы + ХY
22 аутосомы + Х 22 аутосомы + 0
F144 аутосомы + Х0
Для женщин с синдромом Шерешевского-Тернера характерны маленький рост, короткая шея, воронкообразная грудина, бесплодие вследствие недоразвития яичников, слабое развитие половых признаков. 50% больных умственно отсталы или нормальны. Могут быть пороки развития внутренних органов. Дети с синдромом Шерешевского-Тернера рождаются с частотой 0,7 на 1000 новорожденных девочек.
Диагноз ставят при исследовании полового хроматина и на основании результатов цитогенетического анализа.
Аутосомные моносомии среди живорожденных очень редки. Это мозаичные организмы с нормальными клетками. Моносомия касается аутосом 21 и 22. Полные трисомии описаны по большому числу хромосом: 8, 9, 13, 14, 18, 21, 22 и Х. Число Х-хромосом у человека может доходить до 5 с сохранением жизнеспособности.
Изменение числа хромосом вызвано нарушением распределения их по дочерним клеткам во время 1-го или 2-го мейотического деления в гаметогенезе или при первых дроблениях оплодотворенной яйцеклетки.
— при расхождении во время анафазы редуплицированной хромосомы, в результате чего удвоенная хромосома попадает только в одну дочернюю клетку;
— при нарушении конъюгации гомологичных хромосом, что может нарушить правильность расхождения гомологов по дочерним клеткам;
— при отставании хромосом в анафазе при их расхождении в дочерние клетки, что может привести к утрате хромосомы.
При нарушении в двух и более последовательных делениях возникают тетрасомии и другие полисомии.
К полисомиям относятся синдром Клайнфельтера, синдром Тернера и трисомия по Х-хромосоме.
Женщины с кариотипом ХХХ встречаются с частотой 1-1,4 на 1000 родившихся девочек. Для больных с кариотипом ХХХ характерно наличие недоразвитых яичников, матки, бесплодие. Умственное развитие нормальное или в пределах нижней границы нормы. Около 30% женщин сохраняют способность иметь детей.
С увеличением числа Х-хромосом в кариотипе до 4, 5 и более клинические проявления синдрома увеличиваются. Больные не могут иметь детей, умственно более отсталы. При исследовании полового хроматина в ядрах клеток эпителия слизистой оболочки щеки обнаруживают 2 и более телец Барра. Впервые синдром трисомии по Х-хромосоме описал П.Джекобе в 1959 г.
1 Х-хромосома: нормальный мужчина XY или больная женщина ХО (синдром Шерешевского-Тернера)
2 Х-хромосомы: нормальная женщина ХХ или больной мужчина XXY (синдром Клайнфельтера)
3 Х-хромосомы: больная женщина ХХХ или больной мужчина ХХХY (синдром Клайнфельтера)
4 Х-хромосомы: больная женщина(полисомия Х) или больной мужчина XXXY (синдром Клайнфельтера)
При синдроме Клайнфельтера, описанном им в 1942 г., у мужчин в ядрах клеток эпителия слизистой оболочки полости рта обнаружено тельце Барра. В кариотипе 47 хромосом (44+XXY). Частота больных с синдромом Клайнфельтера колеблется в пределах 2-2,5 на 1000 новорожденных мальчиков.
Для мужчин с синдромом Клайнфельтера характерен высокий рост, длинные конечности, евнухоидизм, нарушенный сперматогенез и бесплодие, гинекомастия, повышенное выделение женских гормонов, склонность к ожирению. Иногда наблюдается антисоциальное поведение и алкоголизм. Степень тяжести симптомов пропорциональна числу добавочных Х-хромосом.
Разновидностью синдрома Клайнфельтера является полисомия по хромосоме Y – синдром XYY (47 хромосом). У мужчин с хромосомным набором XYY рост выше среднего, умственное развитие ниже нормы. Они отличаются агрессивным поведением, наблюдается бесплодие. Среди новорожденных мальчики с данным синдромом рождаются с частотой 1:1000.
Полные трисомии описаны по большому числу аутосом: 8, 9, 13,14, 18, 21, 22.
Трисомия по хромосоме 8 приводит к живорождению, но часто наблюдается мозаицизм. Рождение детей с этим геномным нарушением происходит с частотой 1:50000 новорожденных. При синдроме отмечается неглубокая умственная отсталость и физическое недоразвитие. Типичны скелетные аномалии, удлиненное туловище, нарушения речи.
Трисомия по 9-й паре хромосом заканчивается внутриутробной гибелью носителя лишней хромосомы. Продолжительность жизни немногих рожденных детей с такой трисомией-9 составляла 3,5 месяца. Для них характерны внутриутробное недоразвитие, черепно-лицевые пороки, аномалии скелета, пороки сердца, почек и других органов.
Трисомия по 13-й паре хромосом (синдром Патау) — была описана в 1960 г. – встречается с частотой 1:5000-7000.
Для синдрома характерны пороки, лица, внутренних органов (сердца, почек, половых органов), полидактилия. Глухота наблюдается в 80-85% случаев. Имеет место ранняя смертность (в течение года 90% больных).
Трисомии по 14-й паре хромосом описаны для мертворожденных. У живорожденных этой патологии не выявлено.
Наиболее часто встречается трисомия по 21-й паре хромосом (синдром Дауна). Клиническое описание этого синдрома было сделано в 1866 г. Английским врачом Дауном. Мальчики и девочки заболевают одинаково часто. Частота рождения детей с синдромом Дауна – 1:700-800 новорожденных. В большинстве случаев при трисомии в кариотипе 47 хромосом.
Больные с синдромом Дауна небольшого роста, слабоумны, имеют физические пороки. Для них характерны небольшая голова со скошенным затылком, косые глазные щели, эпикант, короткий нос с широкой переносицей, маленькие деформированные уши, полуоткрытый рот с высунутым языком и выступающей челюстью, походка с неловкими движениями, косноязычие. Они имеют пороки сердца, желудочно-кишечного тракта, почек. У больных часто возникают инфекционные и злокачественные заболевания, что обусловлено дефектами иммунной системы. Особенности дерматоглифики связаны с глубокой поперечной бороздой (обезьянья складка) и единственной сгибательной складкой на мизинцах. Благодаря улучшению условий жизни и медицинской помощи, больные с синдромом Дауна доживают до 30 лет и более. Некоторые больные могут заниматься посильной трудовой деятельностью.
Трисомия по 22-й паре, как правило, вызывает летальный эффект и гибель плода во внутриутробном периоде.
Такие генетические нарушения как эуплоидия и анэуплоидия относятся к числу геномных мутаций, т.е. мутаций, связанных с изменением первоначального числа хромосом в клетке.
Необходимо научиться предотвращать данные заболевания во избежание возникновения тяжелых обширных патологий в еще не сформировавшемся человеческом организме. Высокая смертность среди новорожденных с генетическими отклонениями, описанными в данной работе, связана с нарушением развития плода на ранних этапах жизни.
Очень часто большую роль в возникновении мутаций играет человеческий фактор, т.к. загрязнение окружающей среды, нездоровый образ жизни, вредные привычки, такие как курение и алкоголизм пагубно сказываются не только на здоровье человека, но и на здоровье его потомства.
Список использованной литературы
Гинтер Е.К. Медицинская генетика. М.: Медицина, 2008.
Грин Н., Стаут У., Тейлор Д. Биология т. 1-3. М.: Мир, 2009.
В.А. Шевченко, Н.А. Топорнина, Н.С. Стволинская Генетика человека. М.: Владос, 2007.
Горбунова В. Н. Медицинская генетика, 2009
Л.А. Рязанова, Учебное пособие, Практические занятия по основам генетики, 2014
Происхождение мутаций: геномные, хромосомные, генные мутации
Происхождение мутаций: геномные, хромосомные, генные мутации
Геномные мутации. Нерасхождение пары хромосом в ходе мейоза вызывает геномные мутации, например трисомию 21 (синдром Дауна). Геномные мутации приводят к хромосомным анеуплоидиям и бывают наиболее частыми мутациями у человека, с частотой 1 случай нерасхождения на 25-50 мейотических делений клетки.
Это минимальная оценка, поскольку последствия большинства таких мутаций настолько серьезны, что анеуплоидные эмбрионы спонтанно прерываются вскоре после зачатия. Геномные мутации также часто выявляют в клетках опухолей.
Хромосомные мутации
Хромосомные мутации, происходящие с частотой приблизительно одна перестройка на 1700 клеточных делений, случаются значительно реже геномных мутаций. Хотя частоты геномных и хромосомных мутаций могут казаться высокими, эти мутации редко передаются от одного поколения следующему, поскольку они обычно несовместимы с жизнью или нормальной репродукцией. Хромосомные мутации также часто обнаруживают в клетках опухолей.
Генные мутации
Генные мутации, включая замены пар оснований, вставки и делеции, возникают по одному из двух основных механизмов: ошибок в нормальном процессе репликации ДНК или вследствие нарушения репарации ДНК после повреждения. Некоторые мутации происходят спонтанно, другие вызываются физическими или химическими агентами, названными мутагенами, поскольку они существенно повышают частоту мутаций.

Ошибки репликации ДНК. Большинство ошибок репликации быстро удаляются из ДНК и корректируются комплексом ферментов репарации ДНК, сначала опознающим, какая из нитей вновь синтезированной двойной спирали содержит неправильное основание, а затем заменяющим его соответствующим комплементарным основанием.
Репарация ДНК должна быть в высшей степени точным процессом; в противном случае число мутаций в организме было бы недопустимым, и наш вид перестал бы существовать. Фермент ДНК-полимераза точно дублирует двойную спираль благодаря строгому правилу комбинации пар оснований (А с Т, С с G) и молекулярной корректировке.
Всего один неправильный нуклеотид попадает в одну из растущих дочерних нитей на 10 миллионов пар оснований (и это при перемещении вдоль хромосомы человека со скоростью около 50 пар оснований в секунду!). Дополнительная проверка ошибок затем корректирует более 99,9% ошибок репликации ДНК. Таким образом, общий показатель мутаций в результате ошибок репликации имеет в высшей степени низкий уровень 10-10 на пару оснований за одно деление клетки.
Поскольку человеческий диплоидный геном содержит приблизительно 6х109 пары оснований ДНК, репликация ошибок приводит менее чем к одной новой мутации пар оснований на деление клетки.
Репарация повреждений ДНК
Считают, что кроме ошибок репликации, от 10 000 до 1 000 000 нуклеотидов на клетку в день повреждаются спонтанными химическими процессами, такими как, например, деметилирование или деаминирование, реакциями с химическими мутагенами (природными или иными) среды и влиянием ультрафиолетового или ионизирующего излучения.
Некоторые, но не все из этих дефектов могут быть исправлены. Даже если повреждение обнаружено и удалено, система репарации может неточно прочитать комплементарную нить и, как следствие, создать мутацию, вводя неправильные основания. Таким образом, в отличие от изменений ДНК, связанных с репликацией, которые обычно корректируются репарационным механизмом, изменения нуклеотидов, возникающие при репарации поврежденной ДНК, часто приводят к стойким мутациям.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Мутации
Автор статьи — Л.В. Окольнова.
Сразу на ум приходят Люди Х… или Человек — Паук …
Но это в кино, в биологии тоже так, но немного более научно, менее фантастично и более обыденно.
Мута́ция (в переводе — изменение) — устойчивое, передающееся по наследству изменение ДНК, происходящее под влиянием внешних или внутренних изменений.
Мутагенез — процесс появления мутаций.
Обыденность в том, что эти изменения (мутации) происходят в природе и у человека постоянно, почти каждодневно.
В первую очередь, мутации подразделяются на соматические — возникают в клетках тела, и генеративные — появляются только в гаметах.
Соматические мутации
Генеративные мутации
Передаются по наследству.
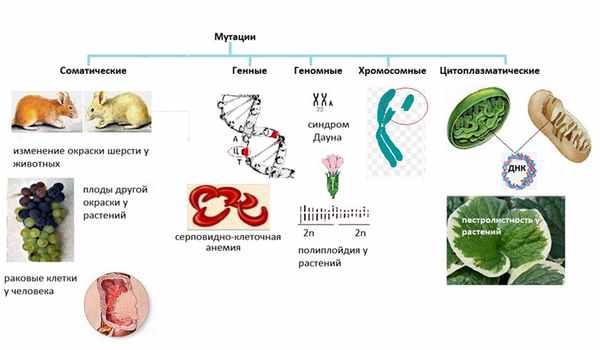
Разберем сначала виды генеративных мутаций.
Генные мутации
Что такое ген? Это участок ДНК (т.е. несколько нуклеотидов), соответственно, это и участок РНК, и участок белка, и какой-либо признак организма.
Т.е. генная мутация — это выпадение, замена, вставка, удвоение, изменение последовательности участков ДНК.
Вообще, это не всегда ведет к болезни. Например, при удвоении ДНК случаются такие “ошибки”. Но они возникают редко, это очень малый процент от всего количества, поэтому они незначительны, что практически не влияют на организм.
Бывают и серьезные мутагенезы:
— серповидно-клеточная анемия у человека;
— фенилкетонурия — нарушение обмена веществ, вызывающее довольно серьезные нарушения умственного развития
— гемофилия
— гигантизм у растений
Геномные мутации
Вот классическое определение термина “геном”:
— совокупность наследственного материала, заключенного в клетке организма;
— геном человека и геномы всех остальных клеточных форм жизни, построены из ДНК;
— совокупность генетического материала гаплоидного набора хромосом данного вида в парах нуклеотидов ДНК на гаплоидный геном.
Для понимания сути мы очень сильно упростим, получится такое определение:
Геном — это количество хромосом
Геномные мутации — изменение числа хромосом организма. В основном, их причина — нестандартное расхождение хромосом в процессе деления.
— синдром Дауна — в норме у человека 46 хромосом (23 пары), однако при этой мутации образуются 47 хромосом
рис. синдром Дауна
— полиплойдия у растений (для растений это вообще норма — большинство культурный растений — полиплойдные мутанты)
Хромосомные мутации — деформации самих хромосом.
Примеры (некоторые перестройки такого рода есть у большинства людей и вообще никак не отражаются ни внешне, ни на здоровье, но есть и неприятные мутации):
— синдром кошачьего крика у ребенка
— задержка в развитии
и т.д.
Цитоплазматические мутации — мутации в ДНК митохондрий и хлоропластов.
Есть 2 органеллы со своими собственными ДНК (кольцевыми, в то время как в ядре — двойная спираль) — митохондрия и растительные пластиды.
Соответственно, есть мутации, вызванные изменениями именно в этих структурах.
Есть интересная особенность — этот вид мутации передается только женским полом, т.к. при образовании зиготы остаются только материнские митохондрии, а “мужские” отваливаются с хвостом при оплодотворении.
Примеры:
— у человека — определенная форма сахарного диабета, туннельное зрение;
— у растений — пестролистность.
Соматические мутации.
Это все описанные выше виды, но возникают они в клетках тела ( в соматических клетках).
Мутантных клеток обычно намного меньше, чем нормальных, и они подавляются здоровыми клетками. (Если не подавляются, то организм перерождаться или болеть).
Примеры:
— у дрозофилы глаз красный, но может иметь белые фасеты
— у растения это может быть целый побег, отличающийся от других (И.В. Мичурин таким образом выводил новые сорта яблок).
— раковые клетки у человека
Примеры вопросов ЕГЭ:
Синдром Дауна является результатом мутации
Генные мутации связаны с изменением
А) числа хромосом в клетках;
Б) структуры хромосом;
B) последовательности генов в аутосоме;
Г) нуклеогидов на участке ДНК.
Мутации, связанные с обменом участками негомологичных хромосом, относят к
Животное, в потомстве которого может появиться признак, обусловленный соматической мутацией
Виды мутаций. Примеры. Причины.

Мутации делятся на несколько основных видов
В биологии все виды мутаций являются одним из эволюционных факторов, так как они могут имеют шанс быть унаследованными потомками мутировавшего организма, микроорганизма или даже клетки, передав эти изменения дальше, что в конечном счёте приведёт к изменению генома. Мутационные процессы, таким образом, оказывают влияние на ход эволюции, причём сами мутации можно разделить на три группы – полезные, нейтральные и вредные. Как показывает изучение закономерностей мутационных процессов, полезных среди них меньше всего, но влияние модификационной изменчивости и этих самых процессов на естественный отбор очень велико.
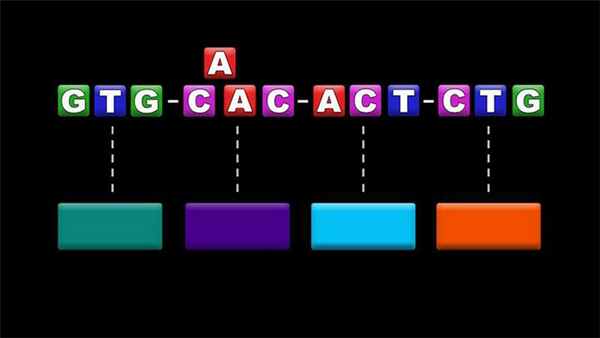
Геномные мутации
Говоря простым языком, это изменение числа хромосом организма. В основном, их причина – нестандартное расхождение хромосом в процессе деления.
- Пример геномной мутации у человека: синдром Дауна, который вызывается наличием “лишней” хромосомы. У человека с этим синдромом хромосом не 46, а 47.
- Пример геномной мутации у растений: полиплойдия. Но в ней нет ничего страшного – например, большинство культурных растений имеют эту мутацию.
Генные мутации
Изменение нуклеотидной последовательности приводит к генной мутации
Это изменение нуклеотидной последовательности в одной молекуле ДНК. При этом в том случае, когда под действием мутации изменяется лишь один нуклеотид, мутация называется точечной. При точечной мутации одно азотистое основание в ДНК или РНК заменяется другим. Если же один участок ДНК заменяется участком другой длины и другого нуклеотидного состава, то такая генная мутация уже называется не точечной, а сложной. Строго говоря, к генной мутации может приводить не только замена одного нуклеотида другим, но и просто вставка другого нуклеотида (без выпадения прежнего) или, наоборот, выпадение нуклеотида без его замены на другой.
Пример генной мутации у растений
- Гигантизм у растений. Возникает он именно из-за генной мутации.
Пример генной мутации у животных и человека
- Фенилкетонурия. Это встречающееся у людей наследственное заболевание, кстати, одно из немногих, поддающихся успешному лечению. Оно связано с нарушением метаболизма аминокислот, главным образом фенилаланина.

Какое влияние оказала научная революция на европейское чудо

30 интересных фактов о Каспийском море
Хромосомные мутации
В отличие от геномных мутаций, они возникают не из-за “неправильного” числа хромосом, а из-за их повреждения. При этом у многих людей можно найти признаки хромосомных мутаций, которые при этом никак не влияют ни на их здоровье, ни на внешность, ни на умственное развитие. Но в некоторых случаях из-за этого мутационного процесса могут возникать хромосомные болезни, которые приводят к наследственным заболеваниям. При этом хромосомные мутации делятся на две категории:
- Внутрихромосомные. Это преобразование генетического материала в пределах одной хромосомы.
- Межхромосомные. Это перестройки, в результате которых две негомологичные хромосомы обмениваются своими участками. Негомологичные хромосомы содержат разные гены и не встречаются в процессе мейоза.
Мутации в соматических клетках
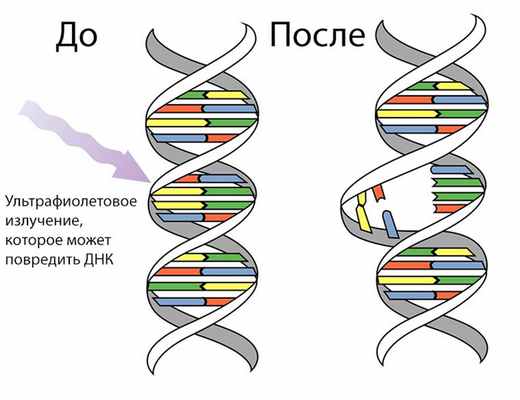
Ультрафиолетовое излучение – одна из причин соматических мутаций
Как мы знаем из биологии, к соматическим клеткам относятся все клетки тела, не имеющие прямого отношения к процессу полового размножения. Соматические мутации возникают в клетках тела и обусловливают мозаичность организма, то есть образование в нём отдельных участков тела, тканей или клеток с отличным от остальных набором хромосом или генов. При этом у них могут возникать мутации всех типов, что и у половых клеток. Одной из их причин, например, могут послужить различные виды излучения. Соматические мутации возникают в соматических клетках, проявляются у данного организма и не передаются потомству при половом размножении. Мутации, возникающие в соматических клетках, наследуются дочерними клетками, которые образуются в процессе митотических делений.
- Пример соматической мутации: окраска шерсти у валлийских овец, у которых присутствует чёрное пятно под хвостом, отличающееся по цвету от остальной части шерстного покрова. Эта мутация могла проявиться либо как доминантная, либо как рецессивная при потере части или всей гомологичной хромосомы.

Образ и характеристика Печорина в «Герое нашего времени»

25 интересных фактов об Эльбрусе
Генеративные мутации
Это мутации, возникающие в клетках полового зачатка и в половых клетках. Собственно, только это и отличает их от соматических мутаций, так как эволюционная ценность генеративных и соматических мутаций различна, она определяется типом размножения организма. При этом генеративные мутации по своей природе ничем не отличаются от соматических.
- Пример генеративной мутации: синдром Дауна у человека. Значительное количество эмбрионов в таких случаях не выживает вовсе, но те из них, что успешно появляются на свет и остаются в живых в первые годы, имеют все шансы дожить до преклонных лет.
Результаты мутаций
Они могут быть самыми разными, как положительными, так и нейтральными или отрицательными. При этом повлиять на дальнейший эволюционный процесс имеют больше шансов именно положительные мутации, так как они предоставляют живым организмам определённые конкурентные преимущества. Отрицательные мутации, которые ослабляют организм тем или иным образом, напротив, снижают шансы организма-мутанта на выживание.
Свойства мутаций

Мутации могут обладать как положительными свойствами, так и негативными
- Они всегда возникают внезапно.
- Они могут передаваться по наследству.
- Из-за отсутствия у них направленного характера их появление невозможно предсказать.
- Они могут быть как полезными, так и вредными или нейтральными.
- Сходные мутации могут возникать многократно.
Причины мутаций

Загрязнение окружающей среды часто приводит к мутациям
Читайте также:
- Синдром Эйзенменгера
- Жгутиконосцы. Трихомоноз. Возбудитель трихомоноза.
- Тромбоз мезентериальных сосудов. Клиника тромбоза мезентериальных сосудов
- Патанатомия несахарного мочеизнурения. Ядра гипоталамуса влияющие на гипофиз
- Лейкодистрофии. Метахроматическая лейкодистрофия
ДНК и гены
ДНК ПРОКАРИОТ И ЭУКАРИОТ
 |
|
, вошедшая в книгу рекордов Гиннесса")
Справа крупнейшая спираль ДНК человека, выстроенная из людей на пляже в Варне (Болгария), вошедшая в книгу рекордов Гиннесса 23 апреля 2016 года
Дезоксирибонуклеиновая кислота. Общие сведения
 Содержание страницы:
Содержание страницы:
- Дезоксирибонуклеиновая кислота
- Строение нуклеиновых кислот
- Репликация
- Строение РНК
- Транскрипция
- Трансляция
- Генетический код
- Геном: гены и хромосомы
- Прокариоты
- Эукариоты
- Строение генов
- Строение генов прокариот
- Строение генов эукариот
- Сравнение строения генов
- Мутации и мутагенез
- Генные мутации
- Хромосомные мутации
- Геномные мутации
- Видео по теме ДНК
- Дополнительный материал
ДНК (дезоксирибонуклеиновая кислота) – своеобразный чертеж жизни, сложный код, в котором заключены данные о наследственной информации. Эта сложная макромолекула способна хранить и передавать наследственную генетическую информацию из поколения в поколение. ДНК определяет такие свойства любого живого организма как наследственность и изменчивость. Закодированная в ней информация задает всю программу развития любого живого организма. Генетически заложенные факторы предопределяют весь ход жизни как человека, так и любого др. организхма. Искусственное или естественное воздействие внешней среды способны лишь в незначительной степени повлиять на общую выраженность отдельных генетических признаков или сказаться на развитии запрограммированных процессов.
Дезоксирибонуклеи́новая кислота (ДНК) — макромолекула (одна из трёх основных, две другие — РНК и белки), обеспечивающая хранение, передачу из поколения в поколение и реализацию генетической программы развития и функционирования живых организмов. ДНК содержит информацию о структуре различных видов РНК и белков.
В клетках эукариот (животных, растений и грибов) ДНК находится в ядре клетки в составе хромосом, а также в некоторых клеточных органоидах (митохондриях и пластидах). В клетках прокариотических организмов (бактерий и архей) кольцевая или линейная молекула ДНК, так называемый нуклеоид, прикреплена изнутри к клеточной мембране. У них и у низших эукариот (например, дрожжей) встречаются также небольшие автономные, преимущественно кольцевые молекулы ДНК, называемые плазмидами.
С химической точки зрения ДНК — это длинная полимерная молекула, состоящая из повторяющихся блоков — нуклеотидов. Каждый нуклеотид состоит из азотистого основания, сахара (дезоксирибозы) и фосфатной группы. Связи между нуклеотидами в цепи образуются за счёт дезоксирибозы (С) и фосфатной (Ф) группы (фосфодиэфирные связи).
 и фосфатной группы.")
Рис. 2. Нуклертид состоит из азотистого основания, сахара (дезоксирибозы) и фосфатной группы
В подавляющем большинстве случаев (кроме некоторых вирусов, содержащих одноцепочечную ДНК) макромолекула ДНК состоит из двух цепей, ориентированных азотистыми основаниями друг к другу. Эта двухцепочечная молекула закручена по винтовой линии.
В ДНК встречается четыре вида азотистых оснований (аденин, гуанин, тимин и цитозин). Азотистые основания одной из цепей соединены с азотистыми основаниями другой цепи водородными связями согласно принципу комплементарности: аденин соединяется только с тимином (А-Т), гуанин — только с цитозином (Г-Ц). Именно эти пары и составляют «перекладины» винтовой «лестницы» ДНК (см.: рис. 2, 3 и 4).

Рис. 2. Азотистые основания
Последовательность нуклеотидов позволяет «кодировать» информацию о различных типах РНК, наиболее важными из которых являются информационные, или матричные (мРНК), рибосомальные (рРНК) и транспортные (тРНК). Все эти типы РНК синтезируются на матрице ДНК за счёт копирования последовательности ДНК в последовательность РНК, синтезируемой в процессе транскрипции, и принимают участие в биосинтезе белков (процессе трансляции). Помимо кодирующих последовательностей, ДНК клеток содержит последовательности, выполняющие регуляторные и структурные функции.

Рис. 3. Репликация ДНК
Расположение базовых комбинаций химических соединений ДНК и количественные соотношения между этими комбинациями обеспечивают кодирование наследственной информации.
Образование новой ДНК (репликация)
- Процесс репликации: раскручивание двойной спирали ДНК — синтез комплементарных цепей ДНК-полимеразой — образование двух молекул ДНК из одной.
- Двойная спираль «расстегивается» на две ветви, когда ферменты разрушают связь между базовыми парами химических соединений.
- Каждая ветвь является элементом новой ДНК. Новые базовые пары соединяются в той же последовательности, что и в родительской ветви.
По завершении дупликации образуются две самостоятельные спирали, созданные из химических соединений родительской ДНК и имеющие с ней одинаковый генетический код. Таким путем ДНК способна перерывать информацию от клетки к клетке.
Более подробная информация:
СТРОЕНИЕ НУКЛЕИНОВЫХ КИСЛОТ

Рис. 4 . Азотистые основания: аденин, гуанин, цитозин, тимин
Дезоксирибонуклеиновая кислота (ДНК) относится к нуклеиновым кислотам. Нуклеиновые кислоты – это класс нерегулярных биополимеров, мономерами которых являются нуклеотиды.
НУКЛЕОТИДЫ состоят из азотистого основания, соединенного с пятиуглеродным углеводом (пентозой) – дезоксирибозой (в случае ДНК) или рибозой (в случае РНК), который соединяется с остатком фосфорной кислоты (H2PO3–).
Азотистые основания бывают двух типов: пиримидиновые основания – урацил (только в РНК), цитозин и тимин, пуриновые основания – аденин и гуанин.

Рис. 5. Структура нуклеотидов (слева), расположение нуклеотида в ДНК (снизу) и типы азотистых оснований (справа): пиримидиновые и пуриновые

Атомы углерода в молекуле пентозы нумеруются числами от 1 до 5. Фосфат соединяется с третьим и пятым атомами углерода. Так нуклеинотиды соединяются в цепь нуклеиновой кислоты. Таким образом, мы можем выделить 3’ и 5’-концы цепи ДНК:

Рис. 6. Выделение 3’ и 5’-концов цепи ДНК
Две цепи ДНК образуют двойную спираль. Эти цепи в спирали сориентированы в противоположных направлениях. В разных цепях ДНК азотистые основания соединены между собой с помощью водородных связей. Аденин всегда соединяется с тимином, а цитозин – с гуанином. Это называется правилом комплементарности (см. принцип комплементарности).
Правило комплементарности:
Например, если нам дана цепь ДНК, имеющая последовательность
3’– ATGTCCTAGCTGCTCG – 5’,
то вторая ей цепь будет комплементарна и направлена в противоположном направлении – от 5’-конца к 3’-концу:
5’– TACAGGATCGACGAGC– 3’.

Рис. 7. Направленность цепей молекулы ДНК и соединение азотистых оснований с помощью водородных связей
РЕПЛИКАЦИЯ ДНК
Репликация ДНК – это процесс удвоения молекулы ДНК путем матричного синтеза. В большинстве случаев естественной репликации ДНК праймером для синтеза ДНК является короткий фрагмент РНК (создаваемый заново). Такой рибонуклеотидный праймер создается ферментом праймазой (ДНК-праймаза у прокариот, ДНК-полимераза у эукариот), и впоследствии заменяется дезоксирибонуклеотидами полимеразой, выполняющей в норме функции репарации (исправления химических повреждений и разрывов в молекле ДНК).
Репликация происходит по полуконсервативному механизму. Это значит, что двойная спираль ДНК расплетается и на каждой из ее цепей по принципу комплементарности достраивается новая цепь. Дочерняя молекула ДНК, таким образом, содержит в себе одну цепь от материнской молекулы и одну вновь синтезированную. Репликация происходит в направлении от 3’ к 5’ концу материнской цепи.

Рис. 8. Репликация (удвоение) молекулы ДНК
ДНК-синтез – это не такой сложный процесс, как может показаться на первый взгляд. Если подумать, то для начала нужно разобраться, что же такое синтез. Это процесс объединения чего-либо в одно целое. Образование новой молекулы ДНК проходит в несколько этапов:
1) ДНК-топоизомераза, располагаясь перед вилкой репликации, разрезает ДНК для того, чтобы облегчить ее расплетание и раскручивание.
2) ДНК-хеликаза вслед за топоизомеразой влияет на процесс «расплетения» спирали ДНК.
3) ДНК-связывающие белки осуществляют связывание нитей ДНК, а также проводят их стабилизацию, не допуская их прилипания друг к другу.
4) ДНК-полимераза δ (дельта), согласовано со скоростью движения репликативной вилки, осуществляет синтез ведущей цепи дочерней ДНК в направлении 5’→3′ на матрице материнской нити ДНК по направлению от ее 3′-конца к 5′-концу (скорость до 100 пар нуклеотидов в секунду). Этим события на данной материнской нити ДНК ограничиваются.

Рис. 9. Схематическое изображение процесса репликации ДНК: (1) Отстающая цепь (запаздывающая нить), (2) Ведущая цепь (лидирующая нить), (3) ДНК-полимераза α (Polα), (4) ДНК-лигаза, (5) РНК-праймер, (6) Праймаза, (7) Фрагмент Оказаки, (8) ДНК-полимераза δ (Polδ), (9) Хеликаза, (10) Однонитевые ДНК-связывающие белки, (11) Топоизомераза.
Далее описан синтез отстающей цепи дочерней ДНК (см. Схему репликативной вилки и функции ферментов репликации)
Нагляднее о репликации ДНК см. видео →
5) Непосредственно сразу после расплетания и стабилизации другой нити материнской молекулы к ней присоединяется ДНК-полимераза α (альфа) и в направлении 5’→3′ синтезирует праймер (РНК-затравку) – последовательность РНК на матрице ДНК длиной от 10 до 200 нуклеотидов. После этого фермент удаляется с нити ДНК.
Вместо ДНК-полимеразы α к 3′-концу праймера присоединяется ДНК-полимераза ε.
6) ДНК-полимераза ε (эпсилон) как бы продолжает удлинять праймер, но в качестве субстрата встраивает дезоксирибонуклеотиды (в количестве 150-200 нуклеотидов). В результате образуется цельная нить из двух частей – РНК (т.е. праймер) и ДНК. ДНК-полимераза ε работает до тех пор, пока не встретит праймер предыдущего фрагмента Оказаки (синтезированный чуть ранее). После этого данный фермент удаляется с цепи.
7) ДНК-полимераза β (бета) встает вместо ДНК-полимеразы ε, движется в том же направлении (5’→3′) и удаляет рибонуклеотиды праймера, одновременно встраивая дезоксирибонуклеотиды на их место. Фермент работает до полного удаления праймера, т.е. пока на его пути не встанет дезоксирибонуклеотид (еще более ранее синтезированный ДНК-полимеразой ε). Связать результат свой работы и впереди стоящую ДНК фермент не в состоянии, поэтому он сходит с цепи.
В результате на матрице материнской нити «лежит» фрагмент дочерней ДНК. Он называется фрагмент Оказаки.
 ДНК-лигаза производит сшивку двух соседних фрагментов Оказаки, т.е. 5′-конца отрезка, синтезированного ДНК-полимеразой ε, и 3′-конца цепи, встроенного ДНК-полимеразой β.
ДНК-лигаза производит сшивку двух соседних фрагментов Оказаки, т.е. 5′-конца отрезка, синтезированного ДНК-полимеразой ε, и 3′-конца цепи, встроенного ДНК-полимеразой β.
СТРОЕНИЕ РНК
Рибонуклеиновая кислота (РНК) — одна из трёх основных макромолекул (две другие — ДНК и белки), которые содержатся в клетках всех живых организмов.
Так же, как ДНК, РНК состоит из длинной цепи, в которой каждое звено называется нуклеотидом. Каждый нуклеотид состоит из азотистого основания, сахара рибозы и фосфатной группы. Однако в отличие от ДНК, РНК обычно имеет не две цепи, а одну. Пентоза в РНК представлена рибозой, а не дезоксирибозой (у рибозы присутствует дополнительная гидроксильная группа на втором атоме углевода). Наконец, ДНК отличается от РНК по составу азотистых оснований: вместо тимина (Т) в РНК представлен урацил (U), который также комплементарен аденину.
Последовательность нуклеотидов позволяет РНК кодировать генетическую информацию. Все клеточные организмы используют РНК (мРНК) для программирования синтеза белков.
Клеточные РНК образуются в ходе процесса, называемого транскрипцией, то есть синтеза РНК на матрице ДНК, осуществляемого специальными ферментами — РНК-полимеразами.
Затем матричные РНК (мРНК) принимают участие в процессе, называемом трансляцией, т.е. синтеза белка на матрице мРНК при участии рибосом. Другие РНК после транскрипции подвергаются химическим модификациям, и после образования вторичной и третичной структур выполняют функции, зависящие от типа РНК.

Рис. 10. Отличие ДНК от РНК по азотистому основанию: вместо тимина (Т) в РНК представлен урацил (U), который также комплементарен аденину.
ТРАНСКРИПЦИЯ
Транскрипция – это процесс синтеза РНК на матрице ДНК. ДНК раскручивается на одном из участков. На одной из цепей содержится информация, которую необходимо скопировать на молекулу РНК – эта цепь называется кодирующей. Вторая цепь ДНК, комплементарная кодирующей, называется матричной. В процессе транскрипции на матричной цепи в направлении 3’ – 5’ (по цепи ДНК) синтезируется комплементарная ей цепь РНК. Таким образом, создается РНК-копия кодирующей цепи.

Рис. 11. Схематическое изображение транскрипции
Например, если нам дана последовательность кодирующей цепи
3’– ATGTCCTAGCTGCTCG – 5’,
то, по правилу комплементарности, матричная цепь будет нести последовательность
5’– TACAGGATCGACGAGC– 3’,
а синтезируемая с нее РНК – последовательность
3’– AUGUCCUAGCUGCUCG – 5’.
ТРАНСЛЯЦИЯ
Рассмотрим механизм синтеза белка на матрице РНК, а также генетический код и его свойства. Также для наглядности по ниже приведенной ссылке рекомендуем посмотреть небольшое видео о процессах транскрипции и трансляции, происходящих в живой клетке:
|
|
В представленном видоролике (кнопка-ссылка слева) показан процесс образования белка из аминокислот. Наглядно (в анимированном варианте) продемонстрированы процессы транскрипции и трансляции. Биосинтез белка на рибосоме также кратко описан в разделе Аминокислоты белков. Более подробное видео о геноме, ДНК и ее структуре, а также процессах кодировки представленно ниже на данной странице: Видео по теме ДНК |


Рис. 12. Процесс синтеза белка: ДНК кодирует РНК, РНК кодирует белок
Трансляция — это процесс, посредством которого генетическая информация преобразуется в белки, рабочие лошадки клетки. Небольшие молекулы, называемые переносными РНК («тРНК»), играют решающую роль в трансляции; они являются молекулами-адаптерами, которые соответствуют кодонам (строительным блокам генетической информации) с аминокислотами (строительными блоками белков). Организмы несут множество типов тРНК, каждая из которых кодируется одним или несколькими генами («набор генов тРНК»).
Вообще говоря, функция набора генов тРНК — переводить 61 тип кодонов в 20 различных типов аминокислот — сохраняется в разных организмах. Тем не менее, состав набора генов тРНК может значительно варьировать между организмами.
ГЕНЕТИЧЕСКИЙ КОД
Генетический код — способ кодирования аминокислотной последовательности белков с помощью последовательности нуклеотидов. Каждая аминокислота кодируется последовательностью из трех нуклеотидов — кодоном или триплетом.
Генетический код, общий для большинства про- и эукариот. В таблице приведены все 64 кодона и указаны соответствующие аминокислоты. Порядок оснований — от 5′ к 3′ концу мРНК.
Таблица 1. Стандартный генетический код
|
1-е ние |
2-е основание |
3-е ние |
|||||||
|
U |
C |
A |
G |
||||||
|
U |
UUU |
Фенилаланин (Phe/F) |
UCU |
Серин (Ser/S) |
UAU |
Тирозин (Tyr/Y) |
UGU |
Цистеин (Cys/C) |
U |
|
UUC |
UCC |
UAC |
UGC |
C |
|||||
|
UUA |
Лейцин (Leu/L) |
UCA |
UAA |
Стоп-кодон** |
UGA |
Стоп-кодон** |
A |
||
|
UUG |
UCG |
UAG |
Стоп-кодон** |
UGG |
Триптофан (Trp/W) |
G |
|||
|
C |
CUU |
CCU |
Пролин (Pro/P) |
CAU |
Гистидин (His/H) |
CGU |
Аргинин (Arg/R) |
U |
|
|
CUC |
CCC |
CAC |
CGC |
C |
|||||
|
CUA |
CCA |
CAA |
Глутамин (Gln/Q) |
CGA |
A |
||||
|
CUG |
CCG |
CAG |
CGG |
G |
|||||
|
A |
AUU |
Изолейцин (Ile/I) |
ACU |
Треонин (Thr/T) |
AAU |
Аспарагин (Asn/N) |
AGU |
Серин (Ser/S) |
U |
|
AUC |
ACC |
AAC |
AGC |
C |
|||||
|
AUA |
ACA |
AAA |
Лизин (Lys/K) |
AGA |
Аргинин (Arg/R) |
A |
|||
|
AUG |
Метионин* (Met/M) |
ACG |
AAG |
AGG |
G |
||||
|
G |
GUU |
Валин (Val/V) |
GCU |
Аланин (Ala/A) |
GAU |
Аспарагиновая кислота (Asp/D) |
GGU |
Глицин (Gly/G) |
U |
|
GUC |
GCC |
GAC |
GGC |
C |
|||||
|
GUA |
GCA |
GAA |
Глутаминовая кислота (Glu/E) |
GGA |
A |
||||
|
GUG |
GCG |
GAG |
GGG |
G |
Среди триплетов есть 4 специальных последовательности, выполняющих функции «знаков препинания»:
- *Триплет AUG, также кодирующий метионин, называется старт-кодоном. С этого кодона начинается синтез молекулы белка. Таким образом, во время синтеза белка, первой аминокислотой в последовательности всегда будет метионин.
- **Триплеты UAA, UAG и UGA называются стоп-кодонами и не кодируют ни одной аминокислоты. На этих последовательностях синтез белка прекращается.
Свойства генетического кода
1. Триплетность. Каждая аминокислота кодируется последовательностью из трех нуклеотидов – триплетом или кодоном.

2. Непрерывность. Между триплетами нет никаких дополнительных нуклеотидов, информация считывается непрерывно.

3. Неперекрываемость. Один нуклеотид не может входить одновременно в два триплета.

4. Однозначность. Один кодон может кодировать только одну аминокислоту.

5. Вырожденность. Одна аминокислота может кодироваться несколькими разными кодонами.

6. Универсальность. Генетический код одинаков для всех живых организмов.
Пример. Нам дана последовательность кодирующей цепи:
3’– CCGATTGCACGTCGATCGTATA– 5’.
Матричная цепь будет иметь последовательность:
5’– GGCTAACGTGCAGCTAGCATAT– 3’.
Теперь «синтезируем» с этой цепи информационную РНК:
3’– CCGAUUGCACGUCGAUCGUAUA– 5’.
Синтез белка идет в направлении 5’ → 3’, следовательно, нам нужно перевернуть последовательность, чтобы «прочитать» генетический код:
5’– AUAUGCUAGCUGCACGUUAGCC– 3’.
Теперь найдем старт-кодон AUG:
5’– AUAUGCUAGCUGCACGUUAGCC– 3’.
Разделим последовательность на триплеты:
![]()
Найдем стоп-кодон и согласно таблице генетического кода запишем последовательность аминокислот:

Центральная догма молекулярной биологии звучит следующим образом: информация с ДНК передается на РНК (транскрипция), с РНК – на белок (трансляция). ДНК также может удваиваться путем репликации, и также возможен процесс обратной транскрипции, когда по матрице РНК синтезируется ДНК, но такой процесс в основном характерен для вирусов.

Рис. 13. Центральная догма молекулярной биологии
ГЕНОМ: ГЕНЫ и ХРОМОСОМЫ
(общие понятия)
Геном — совокупность всех генов организма; его полный хромосомный набор.
Термин «геном» был предложен Г. Винклером в 1920 г. для описания совокупности генов, заключенных в гаплоидном наборе хромосом организмов одного биологического вида. Первоначальный смысл этого термина указывал на то, что понятие генома в отличие от генотипа является генетической характеристикой вида в целом, а не отдельной особи. С развитием молекулярной генетики значение данного термина изменилось. Известно, что ДНК, которая является носителем генетической информации у большинства организмов и, следовательно, составляет основу генома, включает в себя не только гены в современном смысле этого слова. Большая часть ДНК эукариотических клеток представлена некодирующими («избыточными») последовательностями нуклеотидов, которые не заключают в себе информации о белках и нуклеиновых кислотах. Таким образом, основную часть генома любого организма составляет вся ДНК его гаплоидного набора хромосом.
Гены — это участки молекул ДНК, кодирующие полипептиды и молекулы РНК
За последнее столетие наше представление о генах существенно изменилось. Ранее геном называли участок хромосомы, кодирующий или определяющий один признак или фенотипическое (видимое) свойство, например цвет глаз.
|
|
|
Рис. 14. Соответствие между кодирующими участками ДНК, мРНК и аминокислотной последовательностью полипептидной цепи. |

В 1940 г. Джордж Бидл и Эдвард Тейтем предложили молекулярное определение гена. Ученые обрабатывали споры гриба Neurospora crassa рентгеновским излучением и другими агентами, вызывающими изменения в последовательности ДНК (мутации), и обнаружили мутантные штаммы гриба, утратившие некоторые специфические ферменты, что в некоторых случаях приводило к нарушению целого метаболического пути. Бидл и Тейтем пришли к выводу, что ген — это участок генетического материала, который определяет или кодирует один фермент. Так появилась гипотеза «один ген — один фермент». Позднее эта концепция была расширена до определения «один ген — один полипептид», поскольку многие гены кодируют белки, не являющиеся ферментами, а полипептид может оказаться субъединицей сложного белкового комплекса.
На рис. 14 показана схема того, как триплеты нуклеотидов в ДНК определяют полипептид — аминокислотную последовательность белка при посредничестве мРНК. Одна из цепей ДНК играет роль матрицы для синтеза мРНК, нуклеотидные триплеты (кодоны) которой комплементарны триплетам ДНК. У некоторых бактерий и многих эукариот кодирующие последовательности прерываются некодирующими участками(так называемыми интронами).
Современное биохимическое определение гена еще более конкретно. Генами называются все участки ДНК, кодирующие первичную последовательность конечных продуктов, к которым относятся полипептиды или РНК, обладающие структурной или каталитической функцией.
Наряду с генами ДНК содержит и другие последовательности, выполняющие исключительно регуляторную функцию. Регуляторные последовательности могут обозначать начало или конец генов, влиять на транскрипцию или указывать место инициации репликации или рекомбинации. Некоторые гены могут экспрессироваться разными путями, при этом один и тот же участок ДНК служит матрицей для образования разных продуктов.
Мы можем приблизительно рассчитать минимальный размер гена, кодирующего средний белок. Каждая аминокислота в полипептидной цепи кодируется последовательностью из трех нуклеотидов; последовательности этих триплетов (кодонов) соответствуют цепочке аминокислот в полипептиде, который кодируется данным геном. Полипептидная цепь из 350 аминокислотных остатков (цепь средней длины) соответствует последовательности из 1050 п.н. (пар нуклеотидов). Однако многие гены эукариот и некоторые гены прокариот прерываются сегментами ДНК, не несущими информации о белке, и поэтому оказываются значительно длиннее, чем показывает простой расчет.
Сколько генов в одной хромосоме?
 Рис. 15. Вид хромосом в прокаритической (слева) и эукариотической клеках. Гистоны (Histones) — обширный класс ядерных белков, выполняющих две основные функции: они участвуют в упаковке нитей ДНК в ядре и в эпигенетической регуляции таких ядерных процессов, как транскрипция, репликация и репарация.
Рис. 15. Вид хромосом в прокаритической (слева) и эукариотической клеках. Гистоны (Histones) — обширный класс ядерных белков, выполняющих две основные функции: они участвуют в упаковке нитей ДНК в ядре и в эпигенетической регуляции таких ядерных процессов, как транскрипция, репликация и репарация.
ДНК прокариот устроена более просто: их клетки не имеют ядра, поэтому ДНК находится непосредственно в цитоплазме в форме нуклеоида.
 Как известно, бактериальные клетки имеют хромосому в виде нити ДНК, уложенной в компактную структуру – нуклеоид. Хромосома прокариота Escherichia coli, чей геном полностью расшифрован, представляет собой кольцевую молекулу ДНК (на самом деле, это не правильный круг, а скорее петля без начала и конца), состоящую из 4 639 675 п.н. В этой последовательности содержится примерно 4300 генов белков и еще 157 генов стабильных молекул РНК. В геноме человека примерно 3,1 млрд пар нуклеотидов, соответствующих почти 29 000 генам, расположенным на 24 разных хромосомах.
Как известно, бактериальные клетки имеют хромосому в виде нити ДНК, уложенной в компактную структуру – нуклеоид. Хромосома прокариота Escherichia coli, чей геном полностью расшифрован, представляет собой кольцевую молекулу ДНК (на самом деле, это не правильный круг, а скорее петля без начала и конца), состоящую из 4 639 675 п.н. В этой последовательности содержится примерно 4300 генов белков и еще 157 генов стабильных молекул РНК. В геноме человека примерно 3,1 млрд пар нуклеотидов, соответствующих почти 29 000 генам, расположенным на 24 разных хромосомах.
Прокариоты (Бактерии).
 Бактерия E. coli имеет одну двухцепочечную кольцевую молекулу ДНК. Она состоит из 4 639 675 п.н. и достигает в длину примерно 1,7 мм, что превышает длину самой клетки E. coli приблизительно в 850 раз. Помимо крупной кольцевой хромосомы в составе нуклеоида многие бактерии содержат одну или несколько маленьких кольцевых молекул ДНК, свободно располагающихся в цитозоле. Такие внехромосомные элементы называют плазмидами (рис. 16).
Бактерия E. coli имеет одну двухцепочечную кольцевую молекулу ДНК. Она состоит из 4 639 675 п.н. и достигает в длину примерно 1,7 мм, что превышает длину самой клетки E. coli приблизительно в 850 раз. Помимо крупной кольцевой хромосомы в составе нуклеоида многие бактерии содержат одну или несколько маленьких кольцевых молекул ДНК, свободно располагающихся в цитозоле. Такие внехромосомные элементы называют плазмидами (рис. 16).
Большинство плазмид состоит всего из нескольких тысяч пар нуклеотидов, некоторые содержат более 10000 п. н. Они несут генетическую информацию и реплицируются с образованием дочерних плазмид, которые попадают в дочерние клетки в процессе деления родительской клетки. Плазмиды обнаружены не только в бактериях, но также в дрожжах и других грибах. Во многих случаях плазмиды не дают никаких преимуществ клеткам-хозяевам, и их единственная задача — независимое воспроизведение. Однако некоторые плазмиды несут полезные для хозяина гены. Например, содержащиеся в плазмидах гены могут придавать клеткам бактерий устойчивость к антибактериальным агентам. Плазмиды, несущие ген β-лактамазы, обеспечивают устойчивость к β-лактамным антибиотикам, таким как пенициллин и амоксициллин. Плазмиды могут переходить от клеток, устойчивых к антибиотикам, к другим клеткам того же или другого вида бактерий, в результате чего эти клетки также становятся резистентными. Интенсивное применение антибиотиков является мощным селективным фактором, способствующим распространению плазмид, кодирующих устойчивость к антибиотикам (а также транспозонов, которые кодируют аналогичные гены) среди болезнетворных бактерий, и приводит к появлению бактериальных штаммов с устойчивостью к нескольким антибиотикам. Врачи начинают понимать опасность широкого использования антибиотиков и назначают их только в случае острой необходимости. По аналогичным причинам ограничивается широкое использование антибиотиков для лечения сельскохозяйственных животных.
См. также: Равин Н.В., Шестаков С.В. Геном прокариот // Вавиловский журнал генетики и селекции, 2013. Т. 17. № 4/2. С. 972–984.
Эукариоты.
Таблица 2. ДНК, гены и хромосомы некоторых организмов
|
Общая ДНК, п.н. |
Число хромосом* |
Примерное число генов |
|
|
Escherichia coli (бактерия) |
4 639 675 |
1 |
4 435 |
|
Saccharomyces cerevisiae (дрожжи) |
12 080 000 |
16** |
5 860 |
|
Caenorhabditis elegans (нематода) |
90 269 800 |
12*** |
23 000 |
|
Arabidopsis thaliana (растение) |
119 186 200 |
10 |
33 000 |
|
Drosophila melanogaster (плодовая мушка) |
120 367 260 |
18 |
20 000 |
|
Oryza sativa (рис) |
480 000 000 |
24 |
57 000 |
|
Mus musculus (мышь) |
2 634 266 500 |
40 |
27 000 |
|
Homo sapiens (человек) |
3 070 128 600 |
46 |
29 000 |
Примечание. Информация постоянно обновляется; для получения более свежей информации обратитесь к сайтам, посвященным отдельным геномным проектам
*Для всех эукариот, кроме дрожжей, приводится диплоидный набор хромосом. Диплоидный набор хромосом (от греч. diploos- двойной и eidos- вид) – двойной набор хромосом (2n), каждая из которых имеет себе гомологичную.
**Гаплоидный набор. Дикие штаммы дрожжей обычно имеют восемь (октаплоидный) или больше наборов таких хромосом.
***Для самок с двумя Х хромосомами. У самцов есть Х хромосома, но нет Y, т. е. всего 11 хромосом.
В клетке дрожжей, одних из самых маленьких эукариот, в 2,6 раза больше ДНК, чем в клетке E. coli (табл. 2). Клетки плодовой мушки Drosophila, классического объекта генетических исследований, содержат в 35 раз больше ДНК, а клетки человека — примерно в 700 раз больше ДНК, чем клетки E. coli. Многие растения и амфибии содержат еще больше ДНК. Генетический материал клеток эукариот организован в виде хромосом. Диплоидный набор хромосом (2n) зависит от вида организма (табл. 2).
Например, в соматической клетке человека 46 хромосом (рис. 17). Каждая хромосома эукариотической клетки, как показано на рис. 17, а, содержит одну очень крупную двухспиральную молекулу ДНК. Двадцать четыре хромосомы человека (22 парные хромосомы и две половые хромосомы X и Y) различаются по длине более чем в 25 раз. Каждая хромосома эукариот содержит определенный набор генов.

Рис. 17. Хромосомы эукариот. а — пара связанных и конденсированных сестринских хроматид из хромосомы человека. В такой форме эукариотические хромосомы пребывают после репликации и в метафазе в процессе митоза. б — полный набор хромосом из лейкоцита одного из авторов книги. В каждой нормальной соматической клетке человека содержится 46 хромосом.

Размер и функция ДНК как матрицы для хранения и передачи наследственного материала объясняют наличие особых структурных элементов в организации этой молекулы. У высших организмов ДНК распределена между хромосомами.
Совокупность ДНК (хромосом) организма называется геномом. Хромосомы находятся в клеточном ядре и формируют структуру, называемую хроматином. Хроматин представляет собой комплекс ДНК и основных белков (гистонов) в соотношении 1:1. Длину ДНК обычно измеряют числом пар комплементарных нуклеотидов (п.н.). Например, 3-я хромосома человека представляет собой молекулу ДНК размером 160 млн п.н.. Выделенная линеаризованная ДНК размером 3*106 п.н. имеет длину примерно 1 мм, следовательно, линеаризованная молекула 3-й хромосомы человека была бы 5 мм в длину, а ДНК всех 23 хромосом (~3*109 п.н., MR = 1,8*1012) гаплоидной клетки – яйцеклетки или сперматозоида – в линеаризованном виде составляла бы 1 м. За исключением половых клеток, все клетки организма человека (их около 1013) содержат двойной набор хромосом. При клеточном делении все 46 молекул ДНК реплицируются и снова организуются в 46 хромосом.
Если соединить между собой молекулы ДНК человеческого генома (22 хромосомы и хромосомы X и Y или Х и Х), получится последовательность длиной около одного метра. Прим.: У всех млекопитающих и других организмов с гетерогаметным мужским полом, у самок две X-хромосомы (XX), а у самцов — одна X-хромосома и одна Y-хромосома (XY).
Большинство клеток человека диплоидны, поэтому общая длина ДНК таких клеток около 2м. У взрослого человека примерно 1014 клеток, таким образом, общая длина всех молекул ДНК составляет 2・1011 км. Для сравнения, окружность Земли — 4・104 км, а расстояние от Земли до Солнца — 1,5・108 км. Вот как удивительно компактно упакована ДНК в наших клетках!
В клетках эукариот есть и другие органеллы, содержащие ДНК, — это митохондрии и хлоропласты. Выдвигалось множество гипотез относительно происхождения ДНК митохондрий и хлоропластов. Общепризнанная сегодня точка зрения заключается в том, что они представляют собой рудименты хромосом древних бактерий, которые проникли в цитоплазму хозяйских клеток и стали предшественниками этих органелл. Митохондриальная ДНК кодирует митохондриальные тРНК и рРНК, а также несколько митохондриальных белков. Более 95% митохондриальных белков кодируется ядерной ДНК.
СТРОЕНИЕ ГЕНОВ
Рассмотрим строение гена у прокариот и эукариот, их сходства и различия. Несмотря на то, что ген — это участок ДНК, кодирующий всего один белок или РНК, кроме непосредственно кодирующей части, он также включает в себя регуляторные и иные структурные элементы, имеющие разное строение у прокариот и эукариот.
Кодирующая последовательность – основная структурно-функциональная единица гена, именно в ней находятся триплеты нуклеотидов, кодирующие аминокислотную последовательность. Она начинается со старт-кодона и заканчивается стоп-кодоном.
До и после кодирующей последовательности находятся нетранслируемые 5’- и 3’-последовательности. Они выполняют регуляторные и вспомогательные функции, например, обеспечивают посадку рибосомы на и-РНК.
Нетранслируемые и кодирующая последовательности составлют единицу транскрипции – транскрибируемый участок ДНК, то есть участок ДНК, с которого происходит синтез и-РНК.
Терминатор – нетранскрибируемый участок ДНК в конце гена, на котором останавливается синтез РНК.
В начале гена находится регуляторная область, включающая в себя промотор и оператор.
Промотор – последовательность, с которой связывается полимераза в процессе инициации транскрипции. Оператор – это область, с которой могут связываться специальные белки – репрессоры, которые могут уменьшать активность синтеза РНК с этого гена – иначе говоря, уменьшать его экспрессию.
Строение генов у прокариот
Общий план строения генов у прокариот и эукариот не отличается – и те, и другие содержат регуляторную область с промотором и оператором, единицу транскрипции с кодирующей и нетранслируемыми последовательностями и терминатор. Однако организация генов у прокариот и эукариот отличается.

Рис. 18. Схема строения гена у прокариот (бактерий) — изображение увеличивается
В начале и в конце оперона есть единые регуляторные области для нескольких структурных генов. С транскрибируемого участка оперона считывается одна молекула и-РНК, которая содержит несколько кодирующих последовательностей, в каждой из которых есть свой старт- и стоп-кодон. С каждого из таких участков синтезируется один белок. Таким образом, с одной молекулы и-РНК синтезируется несколько молекул белка.
Для прокариот характерно объединение нескольких генов в единую функциональную единицу – оперон. Работу оперона могут регулировать другие гены, которые могут быть заметно удалены от самого оперона – регуляторы. Белок, транслируемый с этого гена называется репрессор. Он связывается с оператором оперона, регулируя экспрессию сразу всех генов, в нем содержащихся.
Для прокариот также характерно явление сопряжения транскрипции и трансляции.

Рис. 19 Явление сопряжения транскрипции и трансляции у прокариот — изображение увеличивается
Такое сопряжение не встречается у эукариот из-за наличия у них ядерной оболочки, отделяющей цитоплазму, где происходит трансляция, от генетического материала, на котором происходит транскрипция. У прокариот во время синтеза РНК на матрице ДНК с синтезируемой молекулой РНК может сразу связываться рибосома. Таким образом, трансляция начинается еще до завершения транскрипции. Более того, с одной молекулой РНК может одновременно связываться несколько рибосом, синтезируя сразу несколько молекул одного белка.
Строение генов у эукариот
Гены и хромосомы эукариот очень сложно организованы
У бактерий многих видов всего одна хромосома, и почти во всех случаях в каждой хромосоме присутствует по одной копии каждого гена. Лишь немногие гены, например гены рРНК, содержатся в нескольких копиях. Гены и регуляторные последовательности составляют практически весь геном прокариот. Более того, почти каждый ген строго соответствует аминокислотной последовательности (или последовательности РНК), которую он кодирует (рис. 14).
Структурная и функциональная организация генов эукариот гораздо сложнее. Исследование хромосом эукариот, а позднее секвенирование полных последовательностей геномов эукариот принесло много сюрпризов. Многие, если не большинство, генов эукариот обладают интересной особенностью: их нуклеотидные последовательности содержат один или несколько участков ДНК, в которых не кодируется аминокислотная последовательность полипептидного продукта. Такие нетранслируемые вставки нарушают прямое соответствие между нуклеотидной последовательностью гена и аминокислотной последовательностью кодируемого полипептида. Эти нетранслируемые сегменты в составе генов называют интронами, или встроенными последовательностями, а кодирующие сегменты — экзонами. У прокариот лишь немногие гены содержат интроны.
Итак, у эукариот практически не встречается объединение генов в опероны, и кодирующая последовательность гена эукариот чаще всего разделена на транслируемые участки – экзоны, и нетранслируемые участки – интроны.
В большинстве случаев функция интронов не установлена. В целом, лишь около 1,5% ДНК человека являются ≪кодирующими≫, т. е. несут информацию о белках или РНК. Однако с учетом крупных интронов получается, что ДНК человека на 30% состоит из генов. Поскольку гены составляют относительно небольшую долю в геноме человека, значительная часть ДНК остается неучтенной.

Рис. 16. Схема строение гена у эукариот — изображение увеличивается
С каждого гена сначала синтезируется незрелая, или пре-РНК, которая содержит в себе как интроны, так и экзоны.
После этого проходит процесс сплайсинга, в результате которого интронные участки вырезаются, и образуется зрелая иРНК, с которой может быть синтезирован белок.

Рис. 20. Процесс альтернативного сплайсинга — изображение увеличивается
Такая организация генов позволяет, например, осуществить процесс альтернативного сплайсинга, когда с одного гена могут быть синтезированы разные формы белка, за счет того, что в процессе сплайсинга экзоны могут сшиваться в разных последовательностях.
Сравнение строения генов прокариот и эукариот

Рис. 21. Отличия в строении генов прокариот и эукариот — изображение увеличивается
МУТАЦИИ И МУТАГЕНЕЗ
Мутацией называется стойкое изменение генотипа, то есть изменение нуклеотидной последовательности.
Процесс, который приводит к возникновению мутаций называется мутагенезом, а организм, все клетки которого несут одну и ту же мутацию — мутантом.
Мутационная теория была впервые сформулирована Гуго де Фризом в 1903 году. Современный ее вариант включает в себя следующие положения:
1. Мутации возникают внезапно, скачкообразно.
2. Мутации передаются из поколения в поколение.
3. Мутации могут быть полезными, вредными или нейтральными, доминантными или рецессивными.
4. Вероятность обнаружения мутаций зависит от числа исследованных особей.
5. Сходные мутации могут возникать повторно.
6. Мутации не направленны.
Мутации могут возникать под действием различных факторов. Различают мутации, возникшие под действием мутагенных воздействий: физических (например, ультрафиолета или радиации), химических (например, колхицина или активных форм кислорода) и биологических (например, вирусов). Также мутации могут быть вызваны ошибками репликации.
В зависимости от условий появления мутации подразделяют на спонтанные — то есть мутации, возникшие в нормальных условиях, и индуцированые — то есть мутации, которые возникли при особых условиях.
Мутации могут возникать не только в ядерной ДНК, но и, например, в ДНК митохондрий или пластид. Соответственно, мы можем выделять ядерные и цитоплазматические мутации.
В результате возникновения мутаций часто могут появляться новые аллели. Если мутантный аллель подавляет действие нормального, мутация называется доминантной. Если нормальный аллель подавляет мутантный, такая мутация называется рецессивной. Большинство мутаций, приводящих к возникновению новых аллелей являются рецессивными.
По эффекту выделяют мутации адаптивные, приводящие к повышению приспособленности организма к среде, нейтральные, не влияющие на выживаемость, вредные, понижающие приспособленность организмов к условиям среды и летальные, приводящие к смерти организма на ранних стадиях развития.
По последствиям выделяются мутации, приводящие к потери функции белка, мутации, приводящие к возникновению у белка новой функции, а также мутации, которые изменяют дозу гена, и, соответственно, дозу белка синтезируемого с него.
Мутация может возникнуть к любой клетке организма. Если мутация возникает в половой клетке, она называется герминативной (герминальной, или генеративной). Такие мутации не проявляются у того организма, у которого они появились, но приводят к появлению мутантов в потомстве и передаются по наследству, поэтому они важны для генетики и эволюции. Если мутация возникает в любой другой клетке, она называется соматической. Такая мутация может в той или иной степени проявляться у того организма, у которого она возникла, например, приводить к образованию раковых опухолей. Однако такая мутация не передается по наследству и не влияет на потомков.
Мутации могут затрагивать разные по размеру участки генома. Выделяют генные, хромосомные и геномные мутации.
Генные мутации
Мутации, которые возникают в масштабе меньшем, чем один ген, называются генными, или точечными (точковыми). Такие мутации приводят к изменению одного и нескольких нуклеотидов в последовательности. Среди генных мутаций выделяют замены, приводящие к замене одного нуклеотида на другой, делеции, приводящие к выпадению одного из нуклеотидов, инсерции, приводящие к добавлению лишнего нуклеотида в последовательность.

Рис. 23. Генные (точечные) мутации
По механизму воздействия на белок, генные мутации делят на: синонимичные, которые (в результате вырожденности генетического кода) не приводят к изменению аминокислотного состава белкового продукта, миссенс-мутации, которые приводят к замене одной аминокислоты на другую и могут влиять на структуру синтезируемого белка, хотя часто они оказываются незначительными, нонсенс-мутации, приводящие к замене кодирующего кодона на стоп-кодон, мутации, приводящие к нарушению сплайсинга:

Рис. 24. Схемы мутаций
Также по механизму воздействия на белок выделяют мутации, приводящие к сдвигу рамки считывания, например, инсерции и делеции. Такие мутации, как и нонсенс-мутации, хоть и возникают в одной точке гена, часто воздействуют на всю структуру белка, что может привести к полному изменению его структуры.
Рис. 25. Схема мутации, приводящей к сдвигу рамки считывания
Хромосомные мутации

Рис. 26. Хромосомные абберации
Хромосомными мутациями называются мутации, которые затрагивают отдельные гены в рамках одной хромосомы. Различают делеции, когда теряется один или несколько генов, дупликации, когда удваивается тот или иной ген или несколько генов, инверсии, когда участок хромосомы поворачивается на 180 градусов, транслокации, когда гены переходят с одной хромосомы на другую.

Рис. 27. Схемы хромосомных мутаций: делеции, дупликации, инверсии
|
|
|
|
Рис. 28. Транслокация |
Рис. 29. Хромосома до и после дупликации |


Геномные мутации
Наконец, геномные мутации затрагивают весь геном целиком, то есть меняется количество хромосом. Выделяют полиплоидии — увеличение плоидности клетки, и анеуплоидии, то есть изменение количества хромосом, например, трисомии (наличие у одной из хромосом дополнительного гомолога) и моносомии (отсутствие у хромосомы гомолога).
Видео по теме ДНК
РЕПЛИКАЦИЯ ДНК, КОДИРОВАНИЕ РНК, СИНТЕЗ БЕЛКА
(Если видео не отображается оно доступно по ссылке→)
См. дополнительно:
- Нуклеиновые кислоты (PDF)
- Общие сведения о секвенировании биополимеров
- Метагеномика и микробиом
- Бактериальный иммунитет и система CRISPR/Cas
- Трансляция белка на рисбосоме (общие сведения)
- Раскрыт секрет спиральной структуры ДНК (новое о ДНК)
- Антимутагенные свойства пробиотиков (в свете защиты ДНК)
- МикроРНК, микробиом кишечника и иммунитет
- Эпигенетика, короткоцепочечные жирные кислоты и врожденная иммунная память
- Замедление старения: роль питательных веществ и микробиоты в модуляции эпигенома (о метилировании ДНК)
Литература в помощь:
Будьте здоровы!
ССЫЛКИ К РАЗДЕЛУ О ПРЕПАРАТАХ ПРОБИОТИКАХ
- ПРОБИОТИКИ
- ПРОБИОТИКИ И ПРЕБИОТИКИ
- СИНБИОТИКИ
- ДОМАШНИЕ ЗАКВАСКИ
- КОНЦЕНТРАТ БИФИДОБАКТЕРИЙ ЖИДКИЙ
- ПРОПИОНИКС
- ЙОДПРОПИОНИКС
- СЕЛЕНПРОПИОНИКС
- ГЕМОПРОПИОВИТ
- БИФИКАРДИО
- ПРОБИОТИКИ С ПНЖК
- МИКРОЭЛЕМЕНТНЫЙ СОСТАВ
- БИФИДОБАКТЕРИИ
- ПРОПИОНОВОКИСЛЫЕ БАКТЕРИИ
- МИКРОБИОМ ЧЕЛОВЕКА
- МИКРОФЛОРА ЖКТ
- ДИСБИОЗ КИШЕЧНИКА
- МИКРОБИОМ и ВЗК
- МИКРОБИОМ И РАК
- МИКРОБИОМ, СЕРДЦЕ И СОСУДЫ
- МИКРОБИОМ И ПЕЧЕНЬ
- МИКРОБИОМ И ПОЧКИ
- МИКРОБИОМ И ЛЕГКИЕ
- МИКРОБИОМ И ПОДЖЕЛУДОЧНАЯ ЖЕЛЕЗА
- МИКРОБИОМ И ЩИТОВИДНАЯ ЖЕЛЕЗА
- МИКРОБИОМ И КОЖНЫЕ БОЛЕЗНИ
- МИКРОБИОМ И КОСТИ
- МИКРОБИОМ И ОЖИРЕНИЕ
- МИКРОБИОМ И САХАРНЫЙ ДИАБЕТ
- МИКРОБИОМ И ФУНКЦИИ МОЗГА
- АНТИОКСИДАНТНЫЕ СВОЙСТВА
- АНТИОКСИДАНТНЫЕ ФЕРМЕНТЫ
- АНТИМУТАГЕННАЯ АКТИВНОСТЬ
- МИКРОБИОМ и ИММУНИТЕТ
- МИКРОБИОМ И АУТОИММУННЫЕ БОЛЕЗНИ
- ПРОБИОТИКИ и ГРУДНЫЕ ДЕТИ
- ПРОБИОТИКИ, БЕРЕМЕННОСТЬ, РОДЫ
- ВИТАМИННЫЙ СИНТЕЗ
- АМИНОКИСЛОТНЫЙ СИНТЕЗ
- АНТИМИКРОБНЫЕ СВОЙСТВА
- КОРОТКОЦЕПОЧЕЧНЫЕ ЖИРНЫЕ КИСЛОТЫ
- СИНТЕЗ БАКТЕРИОЦИНОВ
- АЛИМЕНТАРНЫЕ ЗАБОЛЕВАНИЯ
- МИКРОБИОМ И ПРЕЦИЗИОННОЕ ПИТАНИЕ
- ФУНКЦИОНАЛЬНОЕ ПИТАНИЕ
- ПРОБИОТИКИ ДЛЯ СПОРТСМЕНОВ
- ПРОИЗВОДСТВО ПРОБИОТИКОВ
- ЗАКВАСКИ ДЛЯ ПИЩЕВОЙ ПРОМЫШЛЕННОСТИ
- НОВОСТИ